All published articles of this journal are available on ScienceDirect.

Cardiovascular Dementia - A Different Perspective
Abstract
The number of dementia patients has been growing in recent years and dementia represents a significant threat to aging people all over the world. Recent research has shown that the number of people affected by Alzheimer’s disease (AD) and dementia is growing at an epidemic pace. The rapidly increasing financial and personal costs will affect the world's economies, health care systems, and many families. Researchers are now exploring a possible connection among AD, vascular dementia (VD), diabetes mellitus (type 2, T2DM) and cardiovascular diseases (CD). This correlation may be due to a strong association of cardiovascular risk factors with AD and VD, suggesting that these diseases share some biologic pathways. Since heart failure is associated with an increased risk of AD and VD, keeping the heart healthy may prove to keep the brain healthy as well. The risk for dementia is especially high when diabetes mellitus is comorbid with severe systolic hypertension or heart disease. In addition, the degree of coronary artery disease (CAD) is independently associated with cardinal neuropathological lesions of AD. Thus, the contribution of T2DM and CD to AD and VD implies that cardiovascular therapies may prove useful in preventing AD and dementia.
INTRODUCTION
Recent epidemiologic and clinico-pathologic data suggest overlaps between AD and cerebrovascular lesions that may magnify the effect of mild AD pathology. This interaction may promote the progression of cognitive decline or even precede neuronal damage and dementia. Vascular pathology in the aging brain and in AD includes cerebral amyloid angiopathy (CAA), multiple or recurrent microhaemorrhages and ischaemic infarcts. AD cases with additional minor cerebrovascular lesions are much more likely to have a history of hypertension or infarcts than "pure" AD patients. There is a documented association of low plasma amyloid-beta peptide ratios (Aβ42/Aβ40) with increased imminent risk for mild cognitive impairment and AD [1]. The observation that the nerve growth factor receptor (NGFR, also known as neurotrophic tyrosine kinase receptor 1 (NTRK1) or TRKA) is down-regulated in AD brains led to the suggestion that an imbalance in NGF/NTRK1 signalling might be involved in AD [2-4]. Heart-derived NGF also regulates the survival and maintenance of sympathetic neurons that innervate the heart and is a crucial factor in myocardial ischaemia, a main cause of sudden death in diabetes mellitus [5]. Thus, researchers are currently exploring the possibility of a connection among AD, VD, T2DM and CD [6, 7]. The risk for dementia is especially high when T2DM occurs together with severe systolic hypertension or heart disease. In addition, the degree of CAD is independently associated with the cardinal neuropathological lesions of AD [8]. Consequently, the association of heart failure with an increased risk of AD and VD suggests that the general advice to keep the heart healthy may prove to keep the brain healthy as well [9]. The strong association of cardiovascular risk factors with AD and VD suggests that these diseases share some biologic pathways. The contribution of cardiovascular disease to AD and VD implies that cardiovascular therapies may prove useful for the treatment or prevention of AD and dementia [10-13]. Since NGF has also been discussed as a crucial molecule in diabetic nephropathy [14], it is considered a potential biomarker for AD and neuro-immune diseases [15-17]. This review intends to unravel some potential links among various diseases including AD, VD, T2DM and CD.
ALZHEIMER’S DISEASE
AD is a progressive neurodegenerative disorder that is characterised by two types of proteinaceous deposits in the brain: extracellular amyloid plaques and intracellular neurofibrillary tangles (NFTs). The term “Alzheimer’s disease” was coined after the name of its discoverer, Alois Alzheimer, a German neuropathologist who first described the symptoms of this disease in a 51-year-old patient named “Auguste”. AD is a specific neurodegenerative disease and remains the most common cause of dementia among the elderly. Today, approximately 37 million people worldwide suffer from diverse forms of dementia, and about 18 million of them have AD [18]. The number of patients afflicted with AD is expected to rise and double by the year 2025. AD disrupts communication between brain regions by causing neurons to lose functionality and die [19]. Common symptoms of AD include progressive loss of memory and cognitive dysfunctions such as language deterioration, impaired visuospatial skills, poor judgement, inability to learn new things, indifference, depression, delusions and other manifestations [20]. AD primarily affects the cerebral cortex (grey matter) of the temporal lobe (involved in memory, auditory stimuli and speech), the parietal lobe (involved in orientation, recognition and perception of stimuli) and the frontal lobe (used for reasoning judgment, emotions, movement and speech) of the brain. Deeper structures of the brain, such as the hippocampus (involved in memory and learning processes), and the amygdala (involved in emotional drives), are also affected in AD [21].
AD is classified into two subsets: early onset familial AD (FAD) and late onset sporadic AD (SAD). FAD is very rare and has been found in relatively small numbers of families [22]. Three genes account for the less prevalent FAD: presenilin1 (PS1), PS2 and the amyloid precursor protein (APP) [23]. Mutations in the APP gene, which is located on chromosome 21, have been postulated to play a major role in the aggregation of a clumped sticky protein fragment known as Aβ [24]. The PS1 and PS2 genes are located on chromosomes 14 and 1, respectively. Mutations in the PS1 gene causes enhanced proteolytic processing of APP and lead to the most aggressive form of FAD. Evidence from rodent studies suggests that PS1 mutation results in impaired hippocampus-dependent learning, which also correlates with reduced adult neurogenesis in the dentate gyrus [25].
SAD is the most prevalent form of the disease, affecting about 90% of AD patients. It affects people over the age of 65, including around 50% of all people over the age of 85. SAD is not caused by a specific gene mutation. The most studied gene in SAD is ApoE (polymorphic protein), which is responsible for the production of a protein involved in cholesterol and lipid transport. It exists in three forms, or alleles, named ApoE2, ApoE3, and ApoE4. The ApoE4 allele is a well-established risk factor for SAD [26].
Based on AD research over the past 50 years, three main hypotheses have been discussed. The oldest hypothesis is the “cholinergic hypothesis”, which states that AD begins as a deficiency in the production of acetylcholine (ACh), a vital neurotransmitter [27]. Loss of cholinergic neurons is coupled specifically with typical clinical symptoms such as memory deficits, impaired attention, cognitive decline and reduced learning abilities. The other two hypotheses are called the “Tau-ist” hypothesis and the “ba-ptist” hypothesis. Due to the accumulation of abnormally folded Aβ and Tau proteins in the brains of AD patients, AD has also been called a protein misfolding disease (proteopathy) [28].
Glucose is transported across the blood brain barrier (BBB) into the extracellular space through specific glucose transporters such as GLUT1. Glucose that enters the glycolytic pathway forms Acetyl-CoA. The memory enhancing and memory stabilising neurotransmitter ACh is produced from Acetyl-CoA and choline under the control of insulin. ACh can then bind to nicotinic and muscarinergic ACh receptors that are thus also targets for the treatment of AD [29-31]. In addition, the binding of ACh to its muscarinergic m1 and m3 receptors can then stimulate the formation of soluble APP (sAPP). sAPP in turn can then modulate synaptic plasticity in the hippocampus and promotes dendritic outgrowth [32].
The “Tau-ist” hypothesis revolves around the microtubule-associated protein Tau (MAPT) [33]. Healthy neurons have an internal cytoskeleton, partly made up of structures called microtubules. Microtubules play an important role in nutrient and neurotransmitter transport, structural integrity and cell regulation within neurons. They are mainly present in neuronal axons. The Tau protein helps to stabilise the microtubules through phosphorylation [34, 35]. However, in AD the Tau proteins become hyperphosphorylated and lose the ability to bind to microtubules. Instead, the hyperphosphorylated Tau proteins bind to each other and tie themselves into “knots” known as NFTs [36]. These NFTs destabilise microtubules and collapse the neuronal transport system. Hence, NFT formation results in malfunctions of the microtubule-mediated transport system as well as in communication among neurons, eventually leading to apoptosis [37].
According to the “Ba-ptist” hypothesis, Aβ deposits are the causal factor for the disease. This theory is gaining popularity among scientists because of the recently discovered impaired vascular and cerebrospinal fluid (CSF) transport of Aβ peptides out of the brain and the resulting increase in risk for senile plaque (SP) formation [38, 39]. SPs and NFTs are associated with a loss of neurons and synapses, brain atrophy and dilatation of the lateral ventricles (due to a loss of brain tissue) [40, 41].
APP is a type-I transmembrane protein that has an extracellular N-terminal domain and a short intracellular C-terminal domain. This protein is expressed in many cell types that include neurons in the central nervous system (CNS). APP undergoes non-amyloidogenic cleavage by the α–secretase, releasing soluble sAPPα (Fig. 1) [42]. The release of sAPP from neurons has been implicated in neurite outgrowth and serine protease inhibitor function [43]. A second cleavage by γ–secretase results in the extracellular secretion of small p3 peptides and the intracellular release of the cytoplasmic tail AICD (APP intracellular domain). Once released, AICD is thought to act as a transcriptional activator and to perform other signalling roles. In contrast, the amyloidogenic cleavage of APP is initiated by β-secretase cleavage, a β-site APP-cleaving enzyme (BACE) that releases soluble sAPPβ [44]. The second part of this pathogenic cleavage is mediated by the γ-secretase, generating a 40 amino acid amyloid peptide (Aβ40) that is soluble and innocuous or a 42 amino acid peptide (Aβ42) that clumps to form insoluble amyloid plaques. The non-amyloidogenic pathway probably occurs in the Golgi network and at the cell surface, whereas the amyloidogenic β-secretase pathway is initiated at the endoplasmic reticulum (ER) [45].
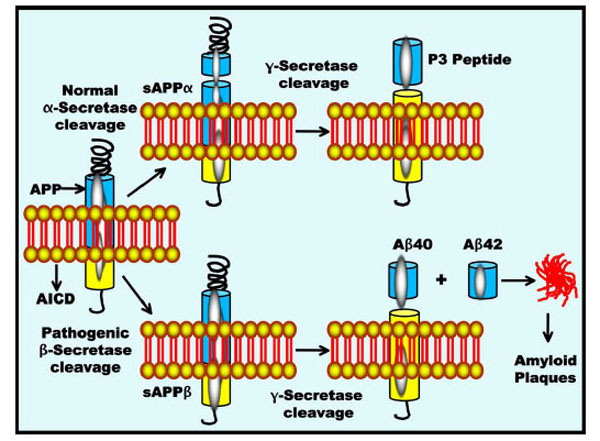
APP processing and Aβ accumulation. APP is metabolised by two competing pathways, the non-pathogenic α-secretase pathway that generates sAPPα and the pathogenic β-secretase pathway that generates sAPPβ. The α-secretase and γ-secretase cleavage releases the p3 peptides, whereas pathogenic β-and γ-secretase cleavage gives rise to Aβ40 and Aβ42 peptide fragments. All carboxy-terminal fragments are substrates for γ-secretase, thus generating the APP intracellular domain (AICD) as well as the secreted peptides p3 and Aβ. Aβ then aggregates into oligomers and subsequently into fibrillary amyloid plaques.
DIABETES
All living organisms need energy to function, and glucose is a potential energy source. The amount of glucose in the bloodstream is regulated by various hormones, the most important of which is insulin. Blood glucose levels are not constant; the level fluctuates depending on the body’s daily needs and hormonal regulations. Under normal conditions, the glucose levels range between 70 and 110 mg/dl. The blood glucose level may rise for three reasons: diet, breakdown of glycogen or hepatic synthesis of glucose. For the human body to function properly, an optimum level of glucose must be maintained in the blood. Extreme fluctuation of the level of glucose in the blood leads to various consequential phenotypes. For instance, diabetes is a metabolic disorder that is categorised into two major subgroups: type-1 diabetes mellitus (T1DM) and type-2 diabetes mellitus (T2DM). This classification is based on whether the elevation of blood glucose level is the result of insulin deficiency (T1DM) or insulin resistance (T2DM) [46]. T1DM is an autoimmune disease that occurs when low levels of insulin are produced by the pancreas due to a malfunction of insulin-producing beta cells. T1DM was previously termed “insulin-dependent diabetes mellitus” (IDDM), or juvenile-onset diabetes, and it accounts for ~5-10% of all diagnosed cases of diabetes. This disorder is lethal unless treatment with an exogenous supply of insulin is used to replace this crucial hormone. T2DM, which was previously termed non-insulin dependent diabetes mellitus (NIDDM), or adult-onset diabetes, accounts for about 90-95% of all diagnosed cases [47, 48]. In T2DM, the pancreas functions normally to produce enough insulin but the cells of the body are resistant to it. T2DM is caused by insulin resistance in peripheral tissues and is most frequently associated with aging, a family history of diabetes, obesity and failure to exercise. Individuals with T2DM have hyperglycaemia and hyperinsulinaemia [49, 50].
GLUCOSE TRIGGERS INSULIN SYNTHESIS AND SIGNALLING
Insulin is a powerful anabolic and anti-catabolic peptide secreted exclusively by beta cells in the islets of Langerhans. These cells accumulate in tiny groups scattered throughout the tissue of the pancreas. Insulin is composed of two chains of amino acids (aa): the 21 aa A chain and the 30 aa B chain. Both chains are linked together by two disulfide bridges. However, a third disulfide bridge exists within the A-chain, linking the 6th and 11th residues of the A-chain. Insulin action is mediated by the IR, a complex multi-subunit cell surface glycoprotein. Binding of insulin activates the intrinsic tyrosine kinase activity of the receptor, which is composed of a pair of α-subunits and a pair of β-subunits. These are distributed mainly in the cortex and hippocampus. Insulin binds to the α-subunits and induces conformational changes that are transmitted to the β-subunits. Subsequently, the β-subunits autophosphorylate and initiate a cascade of phosphorylation and dephosphorylation reactions [51, 52].
Glucose is transported into beta cells by GLUT2 (Type 2 glucose transporters); it then undergoes the universal energy-producing pathway (glycolytic pathway) and enters the respiratory cycle. This produces the high energy ATP molecule via oxidation in the mitochondria. The increase in the ATP:ADP ratio also increases the number of ATP-gated potassium channels (K+) in the beta cell membrane, preventing K+ ions from being shunted across the cell membrane (Fig. 2) [53]. The ensuing rise in positive charge inside the cell, due to increased concentration of K+ ions, leads to a depolarisation of the cell. The net effect is the activation of voltage-gated calcium channels (Ca2+), which transport calcium ions into the cell [54]. The brisk increase in intracellular Ca2+ level causes the activation of phospholipase C (PLC), which cleaves membrane phospholipid phosphatidylinositol-4,5-biphosphate (PIP2) into inositol-1,4,5-triphosphate (IP3) and diacylglycerol (DAG). IP3 then binds to ‘receptor proteins’ in the membrane of the ER. This triggers a rise in the concentration of Ca2+ in the cells and the subsequent activation (together with DAG) of protein kinase C (PKC) and finally the release of pre-synthesised insulin, which is stored in secretory vesicles. The Ca2+ level also regulates the expression of the insulin gene via the cAMP response element binding protein (CREB). This mechanism of glucose action on insulin secretion is similar to that in the pancreas and the brain [55].
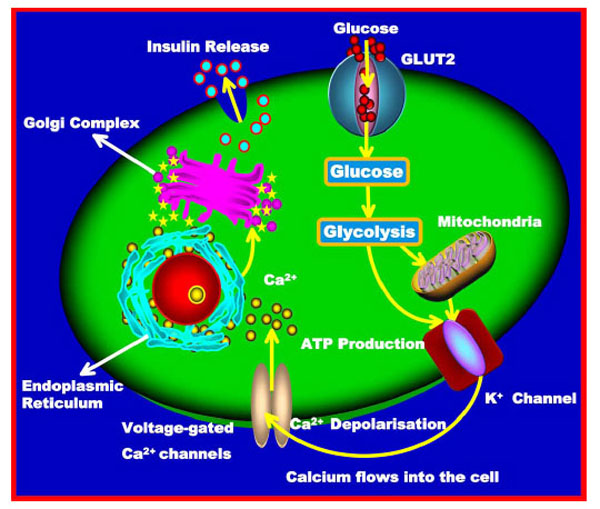
Glucose triggers insulin synthesis and signalling. Insulin secretion in beta cells is triggered by rising blood glucose levels. Starting with the uptake of glucose by the GLUT2 transporter, the glycolytic phosphorylation of glucose causes an increase in the ATP:ADP ratio. This increase inactivates the K+ channel that depolarises the membrane, causing the Ca2+ channel to open up and allowing Ca2+ ions to flow inward. The ensuing increase in levels of Ca2+ ions leads to the exocytotic release of pre-synthesised insulin stored in vesicles. Finally, insulin is released from the storage vesicles.
GLUCOSE AND ITS TRANSPORT IN THE BRAIN
Glucose is an essential fuel for the body and its metabolism is crucial for the brain. Glucose is metabolised by glycolysis and oxidative breakdown in both astrocytes and neurons. Neurovascular and neurometabolic coupling help the brain to maintain an appropriate energy flow to neural tissues under conditions of increased neuronal activity. Glucose in the CNS is transported via facilitated diffusion. This is mediated by a certain glucose transporter family termed “GLUT”. These transporters are localised to the BBB, endothelial cells and cortical membranes. In the brain, GLUT1, GLUT2 and GLUT5 are expressed predominantly in a cell-specific manner [56-58]. Two forms of GLUT1 exist in the brain at different molecular weights (MW): 55 kDa and 45 kDa. The MW depends on the degree of glycosylation of GLUT1. The 55 kDa form is essentially localised to brain microvessels, choroid plexus and vascular endothelial cells, whereas the 45 kDa form is found predominantly in astrocytes [59, 60]. Although GLUT3 is considered specific to neurons, it has also been detected in brain astroglial cells, brain endothelial cells and other tissues such as the placenta, liver and kidney. GLUT5 is localised to microglial cells, the resident macrophages of the brain. It plays a pivotal role in the immune and inflammatory responses of the nervous system and can also be found in the brain endothelium. Another member of the GLUT family, GLUT2, is selectively localised to astrocytes of discrete brain areas such as the hypothalamic and brain stem nuclei. These areas participate in the regulation of feeding behaviour and in the central control of insulin release. Other GLUT members that are selectively dispersed in the brain are the insulin-sensitive GLUT4 and GLUT8. GLUT4 is also found in the pituitary and the hypothalamus. However, GLUT8 is expressed only in the hippocampus, the cerebral cortex and the hypothalamus [61-63]. In actuality, glucose enters the brain through the 55 kDa GLUT1 transporters of the endothelial cells of the BBB. In astrocytes, glucose uptake is mediated by the 45 kDa GLUT1 transporters, whereas GLUT3 transporters mediate this process in neurons. Finally, GLUT5 has been found to mediate the uptake of an unidentified substrate into microglial cells [64].
INSULIN AND ITS RECEPTORS IN THE BRAIN
The most contentious feature of brain insulin research concerns the origin of insulin in the brain. Insulin in the brain basically originates from pancreatic β-cells, but is also partially formed in pyramidal neurons such as those in the hippocampus, the prefrontal cortex, the entorhinal cortex and the olfactory bulb; however, it is not synthesized by glial cells. Insulin can enter circum ventricular regions that lack a BBB, and it can also cross the BBB via insulin transporter-mediated active transport [65]. Insulin gene expression and synthesis have been demonstrated in both mature and immature mammalian neuronal cells, yet the consensus is that little or no insulin is produced in the CNS [66]. Insulin acts as a mediator in the communication between the peripheral endocrine system and the brain via various steps of the neuroendocrine axis. The insulin receptor is a heterotetrameric tyrosine kinase receptor that undergoes dimerisation and tyrosine autophosphorylation upon binding insulin. Insulin receptors are widely distributed in the brain, with the highest densities in the olfactory bulb, the hypothalamus, the cerebral cortex and the hippocampus [67]. In the brain, insulin/insulin-like growth factor 1 (IGF1) and their cognate receptors regulate a multitude of processes such as glucose metabolism, neuronal differentiation, stem cell proliferation, cellular repair mechanisms and cognitive function [68, 69]. Increased concentration of peripheral insulin modulates BBB insulin transporters to regulate its transport into the brain [66, 70]. IGF1 is also synthesised locally in many tissues, including the brain. In contrast to insulin, IGF1 in the circulation and in tissues is bound to high affinity IGF binding proteins (IGFBPs) [71]. The receptor for IGF1 is structurally similar to the insulin receptor; both are tyrosine kinases that phosphorylate similar insulin receptor substrate proteins [72]. Glucose metabolism and insulin receptor mediated signal transduction control the activity of several enzymes involved in the generation of the APP derivative Aβ and the phosphorylation of the Tau protein, the two hallmarks of AD.
INSULIN/IGF1 SIGNALLING PATHWAYS AND AD
Insulin/IGF1 Signalling in APP Processing
Impaired insulin and insulin-like growth factor expression and signaling mechanisms are found in AD and considered as type 3 diabetes [70, 73, 74]. Insulin and IGF1 receptors are tyrosine kinases that mediate the phosphorylation of insulin receptor substrates (IRSs) including IRS1-4. Phosphorylated IRS proteins activate multiple signalling pathways that include PI3K and the extracellular signal-regulated kinase (ERK) cascades, which directly modulate various physiological processes [75]. PI3K is an important mediator of the metabolic signalling pathway. This kinase is a dimer composed of a 110-kDa catalytic subunit and a p55-kDa or p85-kDa regulatory subunit. Tyrosine-phosphorylated IRS associates with the two SH2 domains of the regulatory subunit p85, which in turn binds the catalytic subunit p110. PI3K is insulin regulated and activates protein kinase B (PKB), otherwise known as AKT. Activated PI3K also promotes the growth of cortical pyramidal neurons that play a key role in learning and memory [76, 77]. AKT then regulates the activity of the glycogen synthase kinase-3α/β (GSK-3α/β). GSK-3β contributes to the hyper-phosphorylation of Tau [78] and regulates the production of the APP derivative Aβ, which is generated intracellularly in the ER/Golgi apparatus [79]. GSK-3β is also functionally important for regulating glycogen metabolism, cell cycle kinetics, proliferation, survival and cell migration. AKT inhibits GSK-3β, which promotes cellular metabolism, proliferation and cell survival (Fig. 3). Simultaneous release of GSK-3α may promote the storage and misfolding of APP metabolites. The subsequent accumulation of Aβ42 leads to neurite and neuronal degeneration [79].
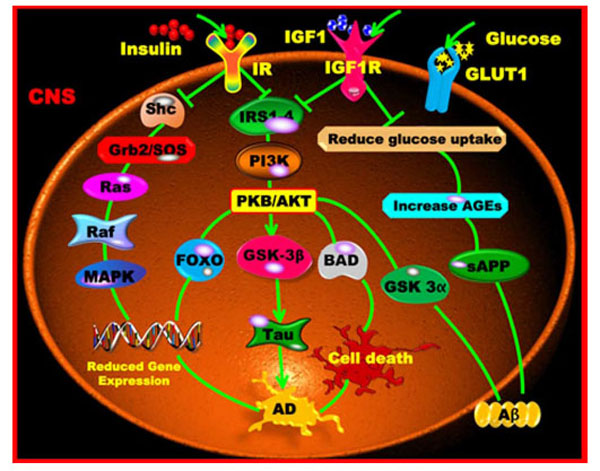
Insulin/IGF1 signalling pathways and AD. Insulin or IGF1 binds with its respective specific receptor sites to activate the IRS proteins that enter the PI3K and PKB/AKT pathways. These pathways cross the GSK-3 and thereby produce the Tau hyper-phosphorylation characteristic for AD. The alternative pathway leads to Shc, Grb2/SOS, Ras, Raf, MAPK activation and gene expression. The absence of IGF1 leads to decreased glucose uptake, decreased glycogen and protein synthesis, an increased level of AGEs, increased Tau phosphorylation and thus increased apoptosis.
Transcription of the genes for insulin and IGF1 receptors is reduced in AD brains compared with controls [80, 81]. AD brains also demonstrate reduced expression of the IRS, IRS-associated PI3K, and activated AKT/PKB [82], suggesting downstream abnormalities in insulin/IGF intracellular signalling mechanisms. As mentioned earlier, activation or inhibition of these signalling mechanisms can influence the metabolism of APP and Aβ. IGF1 also blocks Aβ toxicity in primary cultures of hippocampal neurons. This inhibition of Aβ toxicity occurs through the activation of ERK and AKT/PKB and the prevention of c-Jun N-terminal kinase (JNK) activation in a PI3K-dependent manner [74, 83]. Recent studies provide further support for PI3K as a modulator of APP trafficking and demonstrate that over-activation of the insulin/IGF-1 signaling pathway may result in feedback inhibition of PI3K through IRS1 and reduced APP trafficking and A β secretion [84]. In vivo, insulin/IGF1 signalling is strongly neuroprotective in a variety of AD models. For example, IGF1 protects against mutant APP signalling and chronic subcutaneous infusion of IGF1 has been reported to reduce CNS Aβ levels in aging rats and amyloid plaques in the AD transgenic mouse (Tg2576) model [85-87]. Indeed, IGF1R may activate a common signaling cascade that now seems to link aging to age-associated pathologies of the brain [88]. A disturbed cross talk between IGF1 and AMP-activated protein kinase might be a possible cause that could underlie vascular dysfunctions observed in AD [89].
Insulin/IGF1 Signalling in Tau Phosphorylation
Impaired insulin/IGF1 signalling results in the hyperphosphorylation of Tau due to reduced activation of PI3K/AKT and increased levels of GSK-3β activity (Fig. 3). Tau is normally phosphorylated by GSK-3β and dephosphorylated by the protein phosphatase PP2A [90, 91]. In addition, GSK-3β can be activated through the inhibition of insulin/IGF1 signalling via the Wnt pathway, which is not dependent on PI3K/AKT phosphorylation. Impairment of Wnt signalling mechanisms have been proposed as one key molecular abnormalities in AD [92]. Hyper-phosphorylated Tau fails to be transported into axons and instead accumulates and aggregates in neuronal perikarya, where it promotes oxidative stress that can cause cell death mediated by apoptosis, mitochondrial dysfunction or necrosis. Therefore, the decreased phosphorylation of GSK-3β observed in NIRKO mice results in an increase in GSK-3β activity and may cause elevated Tau phosphorylation [93]. Another study used a mouse model in which the disruption of IR tyrosine kinase signalling led to an accumulation of phosphorylated Tau in the hippocampus. ERK, mitogen-activated protein kinase (MAPK) and cyclin-dependent kinase 5 (CDK-5) are important for the physiological phosphorylation of Tau protein, both of which are activated by insulin/IGF1 receptor. A deficit in insulin concentration increases the activity of GSK-3β, which has been found to cause Tau hyper-phosphorylation and APP metabolism. This may suppress neuronal survival under neurodegenerative conditions [94-96].
DIABETIC COMPLICATIONS AND NEURODEGENERATION
Insulin resistance and hyperinsulinaemia are associated with elevated inflammatory markers and increase the risk of AD [97]. The hallmarks of insulin resistance include selective impairment of the PI3K intracellular signalling pathway in response to insulin, compensatory hyperinsulinaemia, elevated free fatty acids (FFA) and inflammation. PKC and DAG signalling pathways are affected during insulin resistance as well. PKC and DAG are intracellular signalling molecules that regulate vasculature through endothelial permeability and vasodilation. The PKC isozymes are a family of 12 related serine/threonine kinases whose normal function is to activate proteins and lipids that are essential for cell survival. PKC activation is a critical step in the pathway to diabetic microvascular complications [98]. This activation leads to changes in vascular permeability, the extracellular matrix (ECM) synthesis, cytokine synthesis, smooth muscle contraction and angiogenesis, as well as oxidative and osmolar stress, both of which are a consequence of the dysmetabolism of diabetes. Increased DAG and Ca2+ promote the overactivation of PKC-β.
PKC-β over-activation is induced by hyperglycaemia or fatty acids through receptor-mediated activation by PLC. Increased PKC activity and disordered FFA metabolism have been implicated in the over-expression of the angiogenic vascular endothelial growth factor (VEGF), nuclear factor-kappa B (NF-κB), transforming growth factor-β (TGF-β), endothelin-1 (ET-1), and intercellular adhesion molecules (ICAMs). It is also involved in the development of diabetic complications such as retinopathy, nephropathy and CD [99]. PKC-β hyperactivity leads to increased vascular permeability, NO dysregulation, and increased leukocyte adhesion, and it is believed to be involved in cell proliferation, differentiation and apoptosis. Furthermore, PKC-β hyperactivity in neural microvessels causes vasoconstriction. This may lead to decreased blood flow, resulting in hypoxia followed by nerve dysfunction in the brain. Additionally, PKC-β activates MAPK, which subsequently phosphorylates transcription factors that are involved in angiogenesis as well as expression of stress-related genes, JNK and heat shock proteins [100]. VEGF plays a critical role not only in angiogenesis but also in the development of the nervous system [101]. Diabetic animal models have shown high levels of PKC-β in a number of tissues including nerves and endothelium. This may further accelerate the loss of nerve conduction [102].
Elevated blood glucose may be directly toxic to nerves and intravenous infusions of glucose increase neuropathic pain [103]. Hyperglycaemia causes the activation of the polyol pathway, leading to the conversion of glucose into sorbitol by aldose reductase (Fig. 4). The accumulation of the organic osmolyte sorbitol leads to compensatory depletion of other intercellular osmolytes, such as myoinositol and taurine, which in turn leads to reduced Na+/K+ adenosine triphosphatase (ATPase) activity. The polyol pathway also generates fructose, which together with several other compounds such as glucose, glucose-6-phosphate and galactose stimulates the non-enzymatic glycation reaction. This is another metabolic effect of hyperglycaemia. In this process, reduction of sugars forms reversible glycosylation products with proteins, some of which rearrange to form irreversible AGEs [104]. These AGEs can crosslink proteins, including neurofilaments and basement membrane components. Vascular abnormalities observed in patients with diabetic polyneuropathy include basement membrane thickening, endothelial hyperplasia, endothelial dysfunction, increased expression of endothelin and alterations in VEGF expression. Pathologic studies of proximal and distal segments of nerves in diabetic polyneuropathy have shown multifocal fibre loss along the length of the nerves, suggesting ischaemia as a pathogenetic contributor [105]. Vascular and metabolic mechanisms likely interact; hyperglycaemia has a number of deleterious effects on blood vessels [106]. Vessel wall matrix proteins may be affected by non-enzymatic glycation. Hyperglycaemia can lead to increased oxidative stress with the production of superoxides and NO. These substances can interact, leading to protein nitration or nitrosylation, lipid peroxidation, DNA damage and endothelial and neuronal cell death. Superoxides also lead to the activation of PKC, which induces vasoconstriction and reduces neuronal blood flow [107].
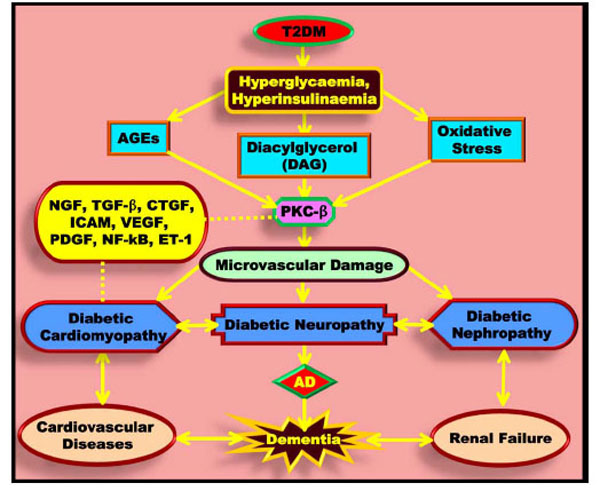
Diabetic complications and neurodegeneration. In diabetes, hyperglycaemia leads to the formation of DAG, AGEs and oxidative stress. These factors enter the PKC-β pathway and produce microvascular damage characteristic of diabetic cardiomyopathy, diabetic neuropathy and diabetic nephropathy.
Diabetic retinopathy (DR) is the most specific microvascular complication and one of the leading causes of visual impairment in diabetic patients. DR complications result from functional and morphological alterations of retinal capillaries, increased permeability to albumin and macromolecules, vascular dysfunction, loss of pericytes and basement membrane thickening. The classic risk factors for diabetic retinopathy are hyperglycaemia, hypertension and hyperlipidaemia. These cardiovascular risk factors are important for both the initial development of diabetic retinopathy and the subsequent progression to advanced retinopathy [108].
Diabetic nephropathy (DN) is a chronic kidney disease characterised by abnormal deposits of matrix material in the glomerular mesangium, leading to glomerulosclerosis. This disorder is also known as intercapillary glomerulonephritis and is described histopathologically by axonal degeneration, demyelination and atrophy, in association with failed axonal regeneration, remyelination and synaptogenesis [109, 110]. DN typically affects the network of tiny blood vessels in the glomerulus, which is necessary for the filtration of blood. The pathophysiology of DN suggests that complex interactions exist between multiple pathways. These include abnormalities in glucose transport mechanisms, increased activity of specific intracellular metabolic pathways, activation of PKC isoforms, formation of reactive oxygen species (ROS) and increased production of AGEs. Altered activity of a variety of growth factors and cytokines may be critical factors in the development of microvascular diabetic complications including nephropathy [111, 112].
ENDOTHELIAL DYSFUNCTION IS A BRIDGE BETWEEN T2DM AND CARDIOVASCULAR DISEASES
The endothelium is a single layer of cells that lines all vessels in the body, including conduit vessels, resistance vessels, pre-capillary arterioles and capillaries. By virtue of its direct contact with the circulating blood, the endothelial layer provides a critical interface between the elements of blood and the tissues. The function of each vessel and the role of its respective endothelium vary according to its location in the body. In the conduit vessels, which include larger arteries such as the aorta and the carotid/coronary/ brachial/femoral arteries, a healthy endothelium provides a smooth, quiescent surface that limits the activation of clotting and proinflammatory factors. This endothelium also blocks the transfer of lipid particles into the arterial wall, inhibits the release of chemokines/cytokines/growth factors and prevents adhesion of platelets and monocytes to the vascular endothelium. In the resistance vessels, endothelial cells (ECs) help to regulate blood flow and systemic blood pressure, whereas in the precapillary arterioles, they play a role in the transport and disposal of waste products of metabolism as well as the distribution of nutrients and hormones such as glucose, fat and insulin. ECs protect blood vessels not only by providing a mechanical lining, but also by controlling vascular tone, inflammation, and thrombosis [113]. EC production of NO and prostacyclin causes vasodilatation, whereas ET-1 mediates vasoconstriction [114]. Inflammatory molecules produced by ECs include interleukin-1 (IL-1), IL-6, IL-18, tumour necrosis factor-α (TNF-α), monocyte chemoattractant protein-1 (MCP-1) and a number of adhesion molecules. Compelling evidence supports endothelial dysfunction as a key early event in the pathogenesis of atherosclerosis. The term endothelial dysfunction encompasses several potential abnormalities; the most extensively studied anomalous characteristic is a reduction in the bioavailability of NO. NO is an antiatherosclerotic signalling molecule released by the vascular endothelium that has potent vasodilatory, anti-inflammatory, anti-proliferative, anti-oxidant, and anti-platelet effects. NO bioavailability is reduced in atherosclerotic vessels before vascular structural changes occur. Impaired NO-dependent vasodilatation is a predictor of future cardiac events and the development of coronary artery atherosclerosis [115-117].
In ECs, insulin increases peripheral vascular blood flow by simulating NO production. FFAs inhibit the insulin-induced increase in peripheral venous NO and peripheral blood flow via two pathways. As illustrated (Fig. 5), the first pathway involves reduction of the tyrosine phosphorylation of IRS-1/2. FFAs inhibit the PI3K/AKT pathway, which, aside from controlling insulin-stimulated glucose uptake, is also needed to stimulate endothelial NOS (e-NOS). Thus, the production of NO in ECs is inhibited. The second inhibitory pathway involves FFA activation of NADPH oxidase in a DAG/PKC-dependent manner. Activation of NADPH oxidase leads to the production of ROS, which results in the destruction of NO [118, 119]. The lipid metabolites activate serine kinases such as PKC and inhibitory κB kinase (IκB), which regulate the activation of nuclear factor-κB (NF-κB), a transcription factor associated with inflammation. The aberrant regulation of NF-κB is associated with a number of chronic diseases including diabetes and atherosclerosis [120]. NF-κB is activated through a common pathway that involves the phosphorylation-induced, proteasome-mediated degradation of the inhibitory subunit IκB. IκB is phosphorylated by the upstream serine kinase IKK, which is phosphorylated and activated by additional upstream serine kinases. Elevated plasma FFA levels also produce low-grade inflammation in skeletal muscle and liver via activation of the NF-κB pathway. This results in the synthesis and release of pro-inflammatory and pro-atherogenic cytokines. Likewise, palmitate activates IKK and JNK, thereby increasing serine phosphorylation of IRS and decreasing insulin stimulated production of NO. In healthy people, insulin infusion results in the release of both NO and ET-1 in adequate concentrations to ensure an appropriate balance for maintaining vascular tone. In contrast, in diabetic patients there is an increased production of ET-1 that creates a vasoconstrictive environment to further impair endothelial function [121]. At the molecular level, insulin exerts its effects through the PI3K or MAPK pathways. In insulin resistance states, the activation of the PI3K pathway is inhibited, whereas the activation of the MAPK pathway is largely intact. The net result is a reduction in vasodilatation and an increase in vasoconstriction, inflammation, and thrombosis that explains much of the increased atherothrombotic risk in diabetic patients [122]. The accumulation of AGE products in diabetes impairs EC function in animal models. Furthermore, AGE products promote vascular inflammation and induce ROS accumulation, and both of these processes contribute to endothelial dysfunction via inhibition of NO production. In addition, acute hyperglycaemia can result in NO reduction and attenuation of endothelium-dependent vasodilatation. Therefore, vascular endothelial dysfunction may occur at any or all levels in the arterial system and contributes to the development and progression of atherosclerosis by favouring coagulation, cell adhesion, and inflammation. This is achieved by promotion of inappropriate vasoconstriction or vasodilation and by enhancement of transendothelial transport of atherogenic lipoproteins [123].

Endothelial dysfunction is a bridge between T2DM and cardiovascular diseases. Increased levels of FFA and DAG trigger a serine/threonine kinase cascade, ultimately inducing serine/threonine phosphorylation of critical IRS-1 sites and thereby inhibiting IRS-1 binding and the activation of PI3-kinase, which results in reduced insulin-stimulated glucose transport.
ROLE OF CYTOKINES IN NEUROINFLAMMATION AND AD
Cytokines also play critical roles in the development and progression of AD. Cells associated with extracellular plaques within the brains of AD patients can produce a variety of cytokines and other related proteins that can ultimately influence plaque and tangle formation. Additionally, Aβ itself can stimulate microglia and astrocytes to secrete pro-inflammatory cytokines, chemokines and reactive ROS, which can lead to neuronal damage [16]. The overproduced cytokines generate a “vicious circle” leading to irreversible synaptic loss and consequently to behaviour alterations. Five major cytokines have been extensively studied in relation to AD onset or progression. Four are pro-inflammatory cytokines: IL-1α, IL-1β, TNF-α and IL-6. TGF-β mainly has an anti-inflammatory role [124]. TNF-α is a critical component of the brain's immune system. Normally, TNF finely regulates the transmission of neural impulses in the brain. Excess TNF-α has been documented in the cerebrospinal fluid of patients with AD [125]. In addition to being secreted by astroglia and microglia, circulating peripheral TNF-α can diffuse through the BBB and access brain cells.
During neuropathologic and neurodegenerative conditions, the level of brain TNF-α increases dramatically, suggesting a significant role of this cytokine in these processes. Whereas the pro-inflammatory effects of TNF-α are most relevant at later stages, TNF-α also has neuroprotective and regenerative functions during the early processes of neurodegenerative diseases [126]. TNF-α has both neurotoxic and neuroprotective effects mediated, respectively, by two receptor subtypes: TNF receptors 1 and 2 (TNFR1 and TNFR2). These two receptors trigger the activation of NF-κB. Most neuroprotective roles of TNF-α are mediated via conscription of NF-κB. In addition to direct IKK mediated activation, NF-κB can also be activated via pro survival kinases such as AKT/PKB. Activated NF-κB dimers enter the nucleus and influence the expression of a complex array of genes constituting the neuronal response to various neurodegenerative disorders. In particular, neuronal NF-κB is proposed to be neuroprotective in nature [127]. In contrast, neurotoxic TNF-α inhibits the transport of Aβ from the brain to the periphery, and elevated TNF-α levels may increase brain accumulation of Aβ [86]. Paradoxically, TNFR1 contains a death receptor domain and has been implicated in pro-apoptotic events, whereas TNFR2 promotes cell survival [128]. Increased levels of TNFR1 and decreased levels of TNFR2 have been observed in AD brains and in peripheral lymphocytes from older adults compared with younger adults though others say that the receptors may not play a major role in the development of AD [129-131]. Aβ42 interacts with inflammatory agents in a cyclically reinforcing manner, such that Aβ elevations increase levels of pro-inflammatory cytokines [132]. In vitro, soluble Aβ oligomers rapidly increase the levels of IL-1β and TNF-α. Conversely, IL-6 and IL-1β can regulate the processing of APP, from which Aβ is derived, and increase the production of Aβ42 [133].
ROLE OF ADVANCED GLYCATION END PRODUCTS (AGEs) IN DIABETIC VASCULAR DISEASES AND AD
Glucose and other reducing sugars react non-enzymatically with protein amino groups to initiate a post-translational modification process known as non-enzymatic glycosylation. This reaction proceeds from reversible Schiff bases to stable, covalently bonded Amadori rearrangement products. Once formed, the Amadori products undergo further chemical rearrangement reactions to form irreversibly bound AGEs, which play a significant role in the pathogenesis of the chronic complications of diabetes mellitus (Fig. 6) [134]. AGEs are a heterogeneous group of structures; those that have already been identified include pyrraline, pentosidine, crossline, and carboxymethyl lysine (CML). These biomolecules are found both inside and outside of cells. Intracellularly, AGEs generally alter functions and cellular transport. However, AGEs that are present extracellularly disrupt cellular adhesion and activate the AGE receptor (RAGE). AGE is used for a broad range of glycation processes (also called the “Maillard reaction”). Hence, hyperglycaemia leads to the formation of AGEs in high concentrations. AGEs have been implicated in the development of oxidative stress, as well as in T2DM. They also play a role in atherosclerosis and renal insufficiency during aging [135]. RAGE is a member of the immunoglobulin superfamily of cell surface molecules and is expressed in a variety of cell types, including ECs, pericytes, mesangial cells, neurons and glia. The receptor-mediated reactions may further contribute to neuronal degeneration. AGE formation causes increased stiffness in the cardiovascular system, leading to high blood pressure and modification of functionally important proteins like tubulin and ATPases. The formation of AGEs is also associated with the increased production of ROS. ROS is also linked to the pathophysiological model of non-enzymatic glycation effects on oxidative stress. The binding of RAGE by AGEs is capable of mediating inflammatory responses and oxidative stress and leads to diabetic vascular complications [136, 137]. AGEs cause many effects, such as blockage of transport through cell membranes, blockage of protein synthesis, reduction of tissue strength and elasticity and fragmentation of molecules that include DNA [138]. AGE immunoreactivity is associated with pathologic hallmarks of AD (NFTs and NPs), cortical neurons, microglia, astrocytes and blood vessels in AD brains [139, 140]. The most prominent species of AGEs in tissues, N-epsilon-CML adducts, are found at their highest levels in the hippocampus. Levels of CML expression in neurons are correlated with NFT pathology. Interestingly, AD patients with T2DM showed enhanced formation of CML, possibly owing to the increased levels of glucose and enhanced oxidative stress [136]. Astrocytes were the major brain cell type shown to contain AGE products by immunohistochemical techniques that involved antibody reaction with AGEs [141]. AGE-immunoreactive astrocytes are also associated with mature amyloid plaques and express elevated neuronal NOS (nNOS). In cultured astrocytes, AGEs increase the release of IL-1β, TNF-α and NO. In a murine microglial cell line, RAGE was shown to be involved in AGE-mediated generation of IL-6, TNF-α and NO. AGEs, in combination with endogenous NO, increased ROS production, resulting in caspase-3-dependent apoptosis in SH-SY5Y neuroblastoma cells [142].
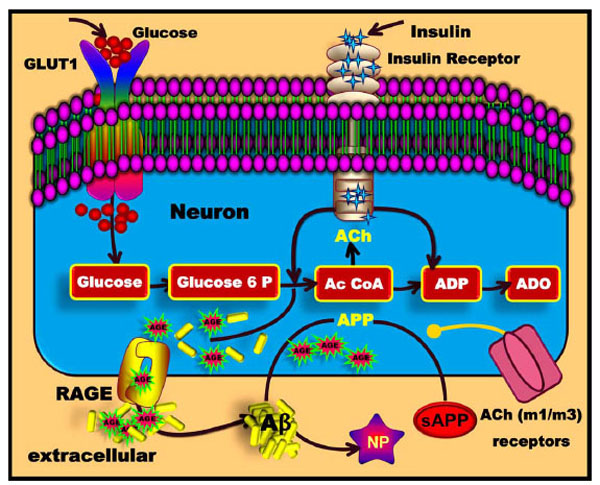
Formation of advanced glycation end products (AGEs) in diabetic vascular diseases and AD. Glucose enters the cell and the glycolytic and tricarboxylic acid (TCA) pathways and eventually produces ATP and amino acids. AGEs are formed from reactions between carbohydrates and proteins. Subsequently, AGEs bind to the RAGE receptors. The RAGE receptor helps Aβ to cross the BBB, after which Aβ forms neuritic plaques (NP).
During normal aging, AGEs accumulate in the brain; this accumulation is implicated in the pathogenesis of AD. AGE products bind to RAGE, which is present in neurons and binds to amyloidogenic APP derivative Aβ [143]. Aβ interaction with RAGE on ECs leads to the transport of Aβ across the BBB and the expression of pro-inflammatory cytokines in those cells [144]. It has also been shown that RAGE stimulates functional BACE1 expression through nuclear factor of activated T-cells 1 (NFAT1) activation, resulting in more Aβ production and deposition in the brain [145]. The deposition of Aβ peptides is thought to be an early and causative event in the pathogenesis of AD and increases markedly during the progression of the disease, leading in turn to the generation of NFTs and finally neuronal death [136, 146].
CHOLESTEROL SYNTHESIS, TRANSPORT AND METABOLISM IN THE BRAIN
Cholesterol is a necessary component of all cell membranes and is important both as a structural component and as a modulator of cell membrane fluidity. Cholesterol is a waxy, fat-like compound that belongs to a class of molecules called steroids. It maintains the integrity of cell membranes and functions to resist changes in temperature and to protect and insulate nerve fibres. It is also involved in the formation of sex hormones (progesterone, testosterone, estradiol, cortisol) and the production of bile salts, which help to digest food. Furthermore, it is converted into vitamin D in the skin when exposed to sunlight. Cholesterol is synthesised in the liver and the brain and can also be obtained through a diet of animal products [147]. It is highly enriched in the brain in comparison with other tissues. In the blood, it is carried on lipoproteins that are classified into chylomicrons, very low-density lipoproteins (VLDL), intermediate-density lipoproteins (IDL), low-density lipoproteins (LDL) and high-density lipoproteins (HDL) [148].
The majority of cholesterol present in the brain is synthesised in the CNS. The brain is the most cholesterol-rich organ in the body and contains about 25% of the total body cholesterol. Cholesterol synthesis begins with the transport of acetyl-CoA from the mitochondria to the cytosol [149]. For every reversal of the thiolase reaction, two molecules of acetyl-CoA are condensed to form acetoacetyl-CoA. Acetoacetyl-CoA and a third molecule of acetyl-CoA are converted to HMG-CoA with the aid of HMG-CoA synthase (Fig. 7). HMG-CoA is converted to mevalonate by HMG-CoA reductase. Mevalonate is then activated by three successive phosphorylations, yielding mevalonate pyrophosphate (MPP). In addition to activating mevalonate, these phosphorylations maintain solubility. After phosphorylation, ATP-dependent decarboxylation yields isopentenyl pyrophosphate (IPP), an activated isoprenoid molecule. IPP is in equilibrium with its isomer, dimethylallyl pyrophosphate (DMPP). One molecule of IPP is condensed with one molecule of DMPP to generate geranyl pyrophosphate (GPP). GPP is further condensed with another IPP molecule to yield farnesyl pyrophosphate (FPP). Thereafter, the NADPH-requiring enzyme squalene synthase catalyses the head-to-tail condensation of two molecules of FPP, yielding squalene (squalene synthase is also tightly associated with the ER). Squalene then undergoes a two step cyclisation to yield inosterol. Finally, the inosterol is converted into cholesterol [147].

Cholesterol synthesis in the brain. Acetyl CoA and acetoacetyl CoA combine to form HMG CoA. The conversion of HMG CoA to mevalonate by HMG CoA reductase is the main regulatory step in the overall pathway. Statins are inhibitors of HMG CoA reductase and inhibit the cholesterol biosynthesis pathway.
Transport of cholesterol from the peripheral circulation into the brain is prevented by an intact BBB. In the CNS, essentially all cholesterol is unesterified and the majority of cholesterol is localised in the myelin sheaths formed by oligodendrocytes. The remaining brain cholesterol is found in neurons, glial cells and extracellular proteins. These pools of cholesterol participate in cholesterol homeostasis of the CNS [150, 151]. Both dietary cholesterol and cholesterol that are synthesised de novo are transported through the circulation in lipoprotein particles [152]. The same is true of cholesterol esters (CEs), the form in which cholesterol is stored in cells. The synthesis and utilisation of cholesterol must be tightly regulated in order to prevent over-accumulation and abnormal deposition within the body. Cholesterol plays an important part in the regulation of Aβ production, with high cholesterol levels being linked to increased Aβ generation and deposition [153]. Abnormalities in the cholesterol metabolism contribute to the development of atherosclerosis, heart disease and AD [154, 155].
Cholesterol is insoluble in the blood and therefore needs carriers known as lipoproteins to aid in its transportation to cells. Astrocytes not only synthesise but also internalise and recycle cholesterol that has been released from degenerating nerve terminals. Astrocytes have the highest capacity to produce ApoE. ApoE binds cholesterol and acts as a ligand for cell surface-lipoprotein receptors such as LDL-receptor-related proteins (LRP), illustrated in (Fig. 8). ApoE is one of the major apolipoproteins found in the plasma. There are spherical and discoidal shaped HDL particles. Under normal conditions, this is an important protein for cholesterol function in the brain and within the CNS; it has the important metabolic function of regulating lipid transport into neurons and clearing cholesterol from ECM [156]. Neurons and glial cells contain LRP receptors, which are large endocytic membrane receptors that can bind and internalise the ApoE-cholesterol phospholipid complex and then fuse with lysosomes. Lysosomes are vesicles that contain the necessary hydrolytic enzymes to allow the intracellular release of free cholesterol (FC). The ATP-binding cassette (ABC) transporter ABCA1 functions to secrete cholesterol from astrocytes and is an important regulator of cholesterol metabolism. In the brain, ABCA1 is also important for cholesterol trafficking, and its expression in cerebral endothelial cells can be stimulated by 24-hydroxycholesterol, thus suggesting a role in the removal of excess brain cholesterol [157].
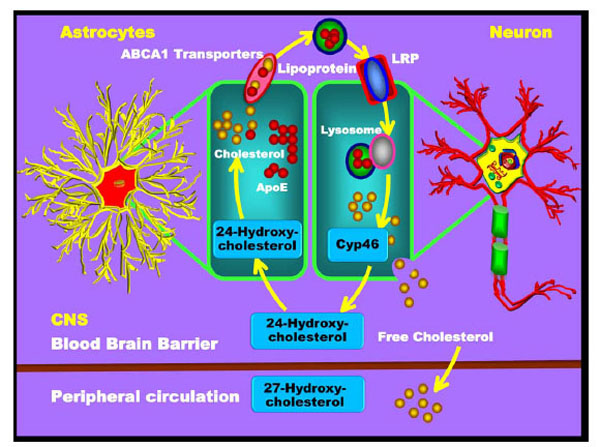
Cholesterol transport and metabolism in the brain. Cholesterol is mainly generated in astrocytes and is then transported to neurons. Cholesterol and ApoE are synthesised by the astrocytes and form the lipoprotein compound. These lipoproteins enter neurons through the neuronal surface receptors of the LDL-receptor related protein (LRP). Upon neuronal entry, the lysosome reacts with the lipoprotein and releases the free cholesterol from the neuron. Some of the free cholesterol reacts with the Cyp46 enzyme and forms the 24-hydroxycholesterol. The 24-hydroxycholesterol is then released from the neuron to enter the astrocytes and to be reconverted into cholesterol. The remaining free cholesterol released from neurons crosses the BBB into the peripheral blood circulation.
ABC transporters, particularly the ABCA1 transporter expressed in astrocytes, may also be of importance for shuttling cholesterol from glial cells to neurons. After cholesterol is synthesised in astrocytes, it is released through ABCA1 and packaged into lipoprotein particles resembling HDL, which in turn are then taken up into neurons through the recognition of ApoE by a variety of lipoprotein receptors such as LDLR, LRP and the ApoE receptor. The cholesterol is then oxidised into 24-hydroxycholesterol and 27-hydroxycholesterol with the help of the enzymes CYP46 and CYP27. The 24-hydroxycholesterol is made by neurons within the brain [158-160]. In contrast, 27-hydroxycholes-terol is produced by many cells, including neurons and oligodendrocytes. Cholesterol 24-hydroxylase, encoded by the CYP46 gene on chromosome 14, is expressed almost exclusively in the brain; only very low levels of its mRNA are found in other tissues such as the liver or testis. In the brain, CYP46 is expressed at especially high levels in neurons found in the cortex, hippocampus, dentate gyrus, thalamus and cerebellar Purkinje cells; however it does not appear to be expressed in glia [161]. CYP46 is a member of the cytochrome P450 family and is responsible for the catabolism of nearly all CNS cholesterol to 24-hydroxycholesterol [162]. A portion of the FC reacts with CYP46 and is converted into 24-hydroxycholesterol in neurons, which can then re-enter the astrocytes for the recycling of cholesterol. The remaining excess cholesterol crosses the BBB and enters into the plasma, where it is eventually delivered to the liver for excretion into bile [163].
APOE AND AD
ApoE protein functions to maintain the synaptic integrity of neurons by stabilising the neuronal cytoskeleton, regulating intracellular Ca2+ levels and regulating the interaction among neurons, ECM and cholesterol. The largest production of ApoE is found in the liver, followed by the brain. In the brain, ApoE is predominantly secreted by astrocytes and microglia [164]. Abundant experimental studies have shown that ApoE influences the majority of the AD pathological processes such as Aβ generation or deposition, NFT formation, neuronal survival, lipid homeostasis and intracellular signalling [165]. There are three common human isoforms, namely ApoE2, ApoE3 and ApoE4, which arise from alleles E2, E3 and E4, respectively. ApoE3 is the most common isoform in the general population (77-78 %), while ApoE2 is found in only 7-8 % and ApoE4 is found in 14-16 % of individuals. ApoE4 is the strongest known risk factor for the development of SAD. The isoforms of ApoE influence Ca2+ channels differentially, resulting in different concentrations of free intracellular Ca2+. ApoE is also involved in various signalling pathways that coincide with the signalling pathways relevant to AD [166]. These pathways also affect the activities of kinases such as PKC, GSK-3β, PKB/AKT, ERK, and JNK, as well as the transcriptional activity of CREB. The ApoE4 allele has also been associated with familial hypercholesterolaemia. The direct effect of hypercholesterolaemia on ApoE levels in the brain leads to an accumulation of Aβ. Neuronal ApoE receptors may also have a number of roles in APP trafficking and processing of Aβ clearance [167]. Moreover, ApoE4 has been found to enhance the synthesis of Aβ by promoting endocytotic recycling of APP. Findings obtained with the transgenic mouse model expressing human APPV717+/- (PDAPP mice) suggest that ApoE4 contributes to the deposition of Aβ [168]. Another in vivo study in transgenic mice showed that carboxy-terminal truncated ApoE stimulates Tau phosphorylation and intracellular formation of NFTs [169].
CHOLESTEROL AND APP PROCESSING
It has been shown that cholesterol accumulates in the dense cores of amyloid plaques. As a component of the cell membrane, cholesterol plays a major role in the metabolism of the single transmembrane protein APP and thus in the production of Aβ [170]. Cholesterol is a major constituent of lipid rafts, together with sphingolipids (sphingomyelin and glycosphingolipids) and glycosylphosphatidylinositol (GPI)-linked proteins. As several transmembrane receptors and cytoplasmic signalling molecules have been shown to concentrate to lipid rafts, they are considered functional hot spots of the plasma membrane. Lipid rafts are assembled in the Golgi complex and are constantly endocytosed. These lipid rafts appear to promote the accumulation of Aβ and the initiation of Aβ aggregation [171, 172]. Increased cholesterol levels may lead to elevated membrane cholesterol, which would cause a change in membrane fluidity and lead to increased APP processing and Aβ production. In vitro studies suggest that increased plasma membrane fluidity may enhance APP/α-secretase interactions and α-secretase enzymatic activity. In contrast, rigid cholesterol-enriched membranes may reduce APP/α-secretase interactions and promote β- and γ- secretase processing. The γ-secretase activity has been identified in cholesterol- and sphingolipidrich membrane microdomains known as lipid rafts. Cholesterol loading increases γ-secretase activity and results in enhanced release of amyloidogenic Aβ in vitro and in vivo. In transgenic mice over-expressing APP, a high cholesterol diet led to an increase in Aβ and NPs and a decrease in sAPPα [173-175].
In addition to the effect of cholesterol on membrane lipid ordering, sub-cellular distribution of cholesterol also regulates the proteolytic processing of APP [176, 177]. Unprocessed, mature APP is transported to the plasma membrane, where it undergoes further cholesterol-dependent processing. Partially matured APP is transported to the Golgi complex, where it completes maturation and is finally transported to the plasma membrane (Fig. 9). APP processing is thought to occur mostly in the trans-Golgi network, the plasma membrane, and the endosomes. In addition, Aβ may be generated in the endo/lysosomal system from endocytosed APP. Intracellular cholesterol plays a critical role in Aβ formation by directing the association of β-secretase with APP. Low intracellular cholesterol favours the processing of APP through the non-amyloidogenic pathway, leading to decreased Aβ deposition. Newly synthesised APP is N-glycosylated in the ER, the site of cholesterol esterification and hydroxylation. β- and γ-secretase activities can be found in newly assembled cholesterol/sphingolipid-rich membrane microdomains in the Golgi [175, 178, 179].

Cholesterol and APP processing. Cholesterol plays a central role in APP metabolism and Aβ synthesis. APP metabolism starts in the ER and then enters the Golgi complex and the plasma membrane. In the plasma membrane, Aβ is released from APP. The free Aβ binds with membrane-associated cholesterol, ultimately resulting in Aβ aggregation. The remaining free Aβ binds with ApoE and the cholesterol complex to re-enter the cell through the LRP receptor system. This ApoE-Aβ complex is degraded by lysosome enzymes to release the free cholesterol and Aβ.
The newly generated Aβ is either released to the extracellular space or remains associated with the plasma membrane and lipid raft structures. Binding of Aβ to the cholesterol/sphingolipid-rich membrane microdomains (lipid rafts) strongly favours Aβ aggregation [180]. Excess unesterified cholesterol in the plasma membrane and ER, as well as mitochondria-derived oxysterols, is loaded onto extracellular lipid acceptors such as ApoE by ABCA1. Monomeric Aβ can bind to numerous molecules in the extracellular space. Binding of ApoE to Aβ regulates not only the aggregation but also the cellular uptake of Aβ. ApoE-Aβ complexes are taken up by the cells through receptor-mediated endocytosis mediated by LRP and LDLR. Lipoprotein-bound cholesteryl esters are hydrolysed in the endo/lysosomal system, and cholesterol is transported through Niemann-Pick disease, type 1 (NPC1)-containing compartment to the ER and Golgi. Endocytosed Aβ also has access to other subcellular compartments through the vesicular transport system [181, 182].
NGF AND THE HEART
Neurotrophins (NTs) are small, highly basic proteins that are essential in the nervous system. They are capable of promoting neuronal cell survival or cell death depending on the environment. Alterations in the NT levels may underlie several medical conditions including pain, depression and obesity. Levels are also altered in disorders of nerve regeneration, learning and memory [183]. NTs have four distinct members: NGF, brain-derived neurotrophic factor (BDNF), NT-3 and NT-4/5. These NTs activate a family of receptor proteins, the TRK receptors. TRKA is primarily a receptor for NGF, TRKB is a receptor for BDNF and NT-4/5, and TRKC is the specific receptor for NT-3. In addition to these specific receptors, all NTs bind to the p75NTR, a member of the TNFR superfamily [184].
NGF was the first nerve growth, survival and differentiation factor discovered, and it plays a crucial role in the sympathetic and sensory nervous systems. It regulates proliferation, differentiation and survival of various cell types of both the CNS and the PNS but also of cells of hematopoietic origin [17]. NGF signalling is mediated via binding to high and low affinity receptors. The high affinity receptor is TRKA, which transmits its signal via an internal tyrosine-kinase domain. The heart is innervated by sympathetic, parasympathetic and sensory nerve fibres expressing TRKA, which, together with the CNS, regulate cardiac functions. The efferent sympathetic and parasympathetic nervous system mediates the opposing neuronal regulation for inotropy and chronotropy of heart performance. NGF plays an important role in the maintenance and survival of such sympathetic neurons and a subset of sensory neurons during and after CNS and PNS development, although the survival of sensory neurons becomes independent of NGF in mature organisms [185].
NGF in the sympathetic nervous system has been considered to function as a target-derived survival factor that is supplied in limited quantities by target organs, such as the heart, to regulate the final number of neurons and density of innervation [186]. In the NGF-null mouse, sympathetic ganglia are not present [187]. Conversely, cardiac-specific NGF over-expression in the mouse causes increased sympathetic innervation density and increased levels of norepinephrine stored in the dysfunctional heart [188, 189]. An influence of NGF on the sympathetic nervous system is also evident in adult animals. Injection of NGF into the left stellate ganglion over several weeks in dogs leads to pronounced sympathetic neuronal growth and nerve sprouting in the heart [190].
Myocardial NGF and sympathetic nerves of the heart appear to be linked in more ways than just neural support provided by the NGF. Once synaptic connections are established, NGF can potentiate synaptic transmission between sympathetic neurons and cardiac myocytes [191]. Modulation of these sympathetic synapses by NGF is acute and reversible and proves that synaptic potentiation works through a presynaptic mechanism. In addition to acute modulatory effects on synaptic efficacy, it has also been demonstrated that NGF produces a long-term enhancement of synaptic transmission of sympathetic neurons and cardiac myocytes [192]. Moreover, there is evidence of feedback inhibition of cardiac myocyte production of NGF at high levels of norepinephrine. The release of NGF by cultured cardiac myocytes is inhibited by norepinephrine, and chronic intravenous infusion of norepinephrine in dogs reduces myocardial NGF content [193]. High sympathetic tone in an innervated organ presumably inhibits its NGF production. This concept has been invoked to explain the concurrence in failing hearts of high sympathetic nerve firing rates, low NGF content and sympathetic neuronal rarefaction. Treatment of co-cultures of sympathetic neurons and cardiac myocytes with NGF, to preferentially activate neuronal TRKA, promotes the release of norepinephrine and increases the myocyte beat rate by fourfold [187]. In contrast, treatment with BDNF activates neuronal p75NTR, inducing the release of ACh and significantly slowing the rate of myocyte contraction [194]. These observations suggest that local secretion of NGF from cardiac myocytes and BDNF from the surrounding vasculature can rapidly exert opposing presynaptic actions to regulate heart rate and contractility. Thus, strategies to interfere with NGF-mediated TRKA activation and BDNF-mediated p75NTR activation may prove beneficial in reducing sympathetic neuron sprouting and norepinephrine release [195-197].
In congestive heart failure, NGF levels in the heart decrease as plasma norepinephrine levels rise; concomitantly, ET-1 plasma levels rise as well. However, it is still not clear how these changes are interrelated. Additionally, a recent report suggests that ET-1 over-expression in the adult mouse heart leads to dilated cardiomyopathy and death [198]. The complex interactions between myocytes and nerves in pathologic states provide insights into the disappointing results of ET-1 blockade in congestive heart failure and yield new pharmacologic targets for the future [199].
Thus, these studies indicate that NGF is involved in the regulation of myocardial performance and contributes to the maintenance of circulatory stability and cardiovascular homeostasis. They also indicate that acute coronary syndrome, atherosclerotic coronary arteries and ischaemic myocardium are characterised by altered circulating levels of NGF [200]. Furthermore, direct gene transfer of NGF into diabetic rat hearts improves silent myocardial ischaemia. Therefore, NGF-TRKA signalling might be a potential therapeutic target for sensory neuropathy in diabetic hearts [201, 202]. Taken together, these results suggest a novel role for NGF as a modulator of cardiac function and a potential link and biomarker for diabetic neuropathy and heart failure.
ROLE OF NGF IN KIDNEY DEVELOPMENT AND DIABETIC NEPHROPATHY
In the development of the kidney, mesenchymal and epithelial interactions play a significant role. Interactions between the epithelial ureter bud and the mesenchyme initiate the development of the metanephric kidney. The mesenchyme stimulates the ureter to grow and branch, allowing differentiation into the collecting ducts. In turn, the ureter induces a part of the mesenchyme to convert into new epithelium. Recently, receptors for growth factors were expressed focally in the interacting epithelium and mesenchyme [203]. NGF is critically involved in kidney development. p75NTR persists at a lower level in adults in glomeruli and a subpopulation of renal interstitial cells. It is expressed during later stages of glomerulogenesis, where it is limited to the mesangium [204]. A recent report showed that NGF and p75NTR are over-expressed in the kidneys of patients affected by various renal disorders, and it was hypothesised that this growth factor may be involved in the pathophysiology of inflammatory kidney diseases including diabetic nephropathy [14, 205]. This hypothesis is also supported by the observation that NGF and its receptors are markedly expressed in renal disorders that are characterised by the enhanced presence of mast cells, macrophages and lymphocytes, cells that are known to produce and release NGF [17, 206].
THE CONNECTION AMONG T2DM, CD, AD AND DEMENTIA
Risk factors such as hypertension, T2DM and hypercholesterolaemia increase the risk of both VD and AD [207, 208]. VD is caused by decreased or interrupted circulation of the blood flow to parts of the brain, and it is predominantly caused by cerebrovascular diseases (CVD), CD or stroke. Among the cardiovascular risk factors related to VD are hypertension, atherosclerosis, CAD, T2DM and smoking. Risk factors for stroke do not independently contribute to dementia, but commonly occur in clusters of risk factors [209].
T2DM also contributes to microvascular changes in the brain, which in turn contribute to cognitive dysfunction and stroke [210-214]. Hyperglycaemia is generally accepted to be the major cause of diabetic microvascular complications and may play an important role in the development of macrovascular diseases. This condition allows cholesterol to build up in the walls of the blood vessels to form plaques that inhibit the heart-mediated oxygen and nutrition supply to various tissues and organs, including the brain [215]. The Framingham study also showed that the risk of developing congestive heart failure was increased in patients with T2DM and that the relative impact of diabetes on congestive heart failure was even greater than that for coronary heart disease [216].
Two major types of heart and blood vessel diseases common in people with diabetes are CAD and CVD. CAD is also called ischemic heart disease and is caused by the hardening or thickening of the blood vessel walls. The blood vessels are blocked by fatty deposits that reduce the blood supply to the heart, resulting in a heart attack. In contrast, CD affects blood flow to the brain, leading to the development of CVD and strokes. Brain cells are then deprived of oxygen and subsequently undergo apoptosis [217]. This decreased blood flow to the brain increases the risk of micro- and macrovascular diseases, which ultimately lead to dementia and stroke [218].
Hypertension is also associated with brain atrophy, large and small artery infarction and white matter lesion progression due to altered auto-regulation of blood flow to the brain. In ischaemic white matter lesions, long-term hypertension causes a narrowing of the lumen of the small arteries and arterioles that nourish the deep white matter. These vascular factors could directly affect the development of AD by causing neuronal death and the accumulation of plaques and tangles [219]. Small artery disease causes periventricular white matter ischaemia and lacunar strokes that manifest as VD [220].
Diabetes and ischaemic heart disease seem to enhance myocardial dysfunction, which leads to accelerated heart failure (diabetic cardiomyopathy). Thus, patients with diabetes are unusually prone to congestive heart failure. Several factors that probably underlie diabetic cardiomyopathy include severe coronary atherosclerosis, prolonged hypertension, chronic hyperglycaemia, microvascular disease, glycosylation of myocardial proteins and autonomic neuropathy [221]. Atherosclerosis is the major cause of dementia and increases the risk of AD. Thus, patients with diabetes often develop CD and CVD that in turn result in atherosclerosis which induces cerebral hypoperfusion, thus leading to cerebral hypoxia. These conditions destabilise synaptic connections and neuronal activity, gradually evolving into a neurodegenerative process characterised by the formation of SPs, NFTs and amyloid angiopathy as observed in VD or AD (Fig. 10) [222, 223].
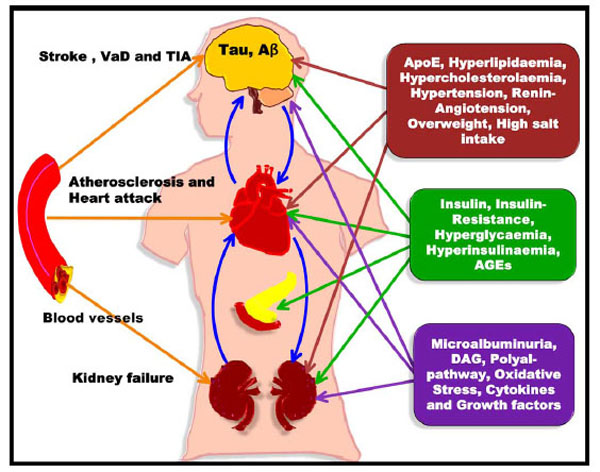
Connection of T2DM, CAD, AD and Dementia. Blockage of heart vessels leads to atherosclerosis and heart attack, whereas blockage of the brain arteries results in VD and stroke.
FUTURE DIRECTIONS AND TREATMENTS OF AD
Insulin-Degrading Enzyme (IDE) as a Therapeutic Target for the Treatment of AD
Insulin is catabolised by IDE, which also degrades other compounds such as IGFI, IGFII, amylin, intra- and extracellular Aβ, glucagon, β-endorphin and dynorphins as well as the AICD fragment produced by the γ-secretase cleavage of APP [224, 225]. IDE is abundant in the cytosol and in peroxisomes. In addition, IDE is found in the rough ER (RER), the plasma membrane, and the extracellular environment. Increased brain levels of insulin should effectively inhibit the degradation of Aβ by IDE. This would consequently enhance Aβ neurotoxicity and aid in AD progression [226]. IDE is part of the negative feedback loops that control insulin action. LRP and RAGE are involved in receptor-mediated flux of Aβ across the BBB (Fig. 11). As mentioned earlier, LRP is a multifunctional signalling and scavenger receptor that consists of a heavy chain capable of binding a variety of ligands such as ApoE and α2-macroglobulin (α2M). α2M is a protein that is capable of binding Aβ with high specificity and thus preventing its fibrillisation [227]. α2M is found in neuritic plaques in AD brains and plays a major role in Aβ clearance via LRP [228]. Aβ can be transported across the BBB and exported out of the brain into the bloodstream either by directly binding to LRP or by binding to the LRP ligands ApoE and α2M. While LRP appears to mediate the efflux of Aβ from the brain to the periphery, RAGE has been strongly implicated in Aβ influx into the CNS. Thus, soluble Aβ can be removed from the brain via two fundamental pathways: either by enzymatic degradation (for instance via IDE) or by receptor-mediated clearance via LRP [229]. Once Aβ crosses the BBB and enters the bloodstream, it can re-enter the brain via the RAGE receptor or be delivered to peripheral sites for degradation in the liver or kidney. Consequently, clearance of Aβ from the brain is a therapeutic approach and can be considered for the prevention of AD [230-232].
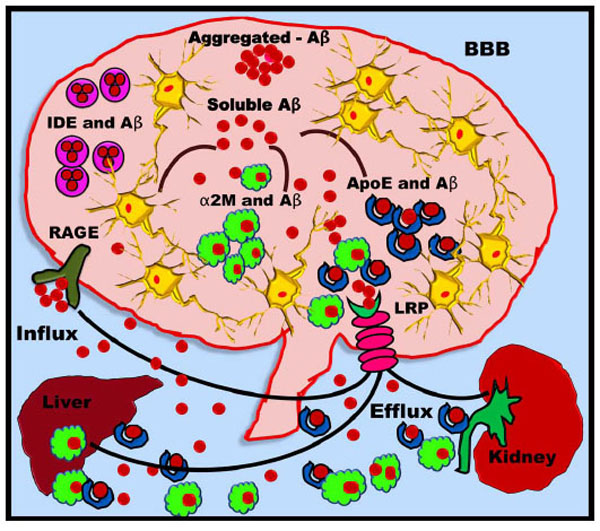
IDE as a therapeutic approach for AD. Aβ is degraded by ApoE, IDE and α2M. Aβ binds to IDE, which allows its degradation inside the CNS. The remaining Aβ binds ApoE and α2M that pass through the BBB via LRP. A portion of soluble Aβ can also efflux from the LRP to re-enter the brain via RAGE. The peripheral ApoE-Aβ and α2M-Aβ complexes enter the liver and kidney, where they are degraded.

AD is classified into two types, FAD and SAD. FAD is based on mutations in specific genes such as PS1, PS2 and APP, whereas SAD is based on various factors including hypercholesterolaemia, hyperglycaemia, and insulin resistance. These SAD factors lead to the formation of T2DM, cardiovascular diseases, kidney diseases, and ultimately neurovascular dementia, such as AD.
ACYL-COA: CHOLESTEROL ACYLTRANSFERASE (ACAT) AS A THERAPEUTIC APPROACH FOR AD
ACAT is a protein that converts cholesterol to highly insoluble CEs. ACAT is essential for the regulation of intracellular cholesterol homoeostasis and the distribution of cholesterol throughout the body. ACAT is the enzyme that removes excess cholesterol from membranes and converts FC to CEs for storage purposes. ACAT has two different isoforms: ACAT1 and ACAT2. In human tissues, ACAT1 is expressed ubiquitously, with prominent protein expression in Kupffer cells (located in the liver) and macrophages, as well as neurons [233]. Expression of ACAT2 appears to be restricted to the liver and intestine, where it is hypothesised to be a major player in the responsiveness to dietary cholesterol. Elevated FC results in the activation of ACAT and the production of CE. A close relationship exists among ACAT, intracellular cholesterol trafficking, and the sterol regulatory element binding protein (SREBP) pathway. SREBP consists of a complex group of membrane proteins that regulate cholesterol homeostasis. ACAT keeps the levels of FC in the membrane under strict control [234]. This pool of FC ultimately regulates the SREBP pathway. In the brain, cholesterol can be either stored within the cell as FC or converted into CEs and stored in cytoplasmic droplets. An increase in the intracellular levels of CE leads to increased Aβ production, whereas inhibition of ACAT leads to decreased levels of CEs and increased intracellular FC. The latter sequence eventually leads to decreased Aβ production. Thus, ACAT is also particularly important in the process of AD development. Pharmacological inhibitors of ACAT are available, and these inhibitors could reduce Aβ production to decrease amyloid plaque formation in AD [235, 236].
STATIN AS A THERAPEUTIC APPROACH FOR AD
Cholesterol synthesis in neurons is regulated by HMG-CoA reductase. HMG-CoA reductase inhibitors are commonly known as statins. Different types of available statins include lovastatin (Mevacer), pravastatin (Prasiocor), simvastatin (Zocor), rosuvastatin (Crestor) and atorvastatin (Lipitor) [237, 238]. These statins competitively inhibit HMG-CoA reductase, which is a rate-limiting enzyme that catalyses the conversion of HMG-CoA to L-mevalonate, and reduce the formation of L-mevalonate. The latter is a key intermediate in cholesterol synthesis and aids in decreasing LDL cholesterol and increasing HDL cholesterol levels [239, 240]. Statins inhibit both cholesterol and isoprenoid synthesis; therefore, it is important to determine their individual effects on APP processing [155, 241, 242]. The variable side chain of a statin determines its solubility across the BBB. Atorvastatin, lovastatin and simvastatin are lipophilic and cross into the brain readily, whereas cerivastatin (Baycol, Lipobay (removed from market)), fluvastatin (Lescol, Canef, Vastin) and pravastatin are hydrophilic and do not cross. Statin inhibitors also inhibit isoprenylation, which results in a reduction of membrane raft clustering involved in APP processing and Aβ degradation [243, 244].
CONCLUSION
The parallel development of T2DM and CD is a compound risk factor for both VD and AD [245]. There are strong additional interactions with other risk factors, such as hypertension, hyperglycaemia and hyperinsulinaemia, hypercholesterolaemia, hyperlipidaemia, endothelial dysfunction, AGE formation and the ApoE4 genotype. A specific role of NGF as potential link (biomarker?) of these diseases, however, remains elusive [15, 17, 246]. Disturbances in insulin signalling pathways may contribute to cognitive decline such as that seen with AD [247]. The link among insulin resistance, cholesterol metabolism, CD and VD has opened a new arena for research in the field of cognitive functions affected by aging and related neurodegenerative diseases.
ABBREVIATIONS
REFERENCES
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link] [PMC Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]
[PubMed Link]

