All published articles of this journal are available on ScienceDirect.

Molecular Anti-diabetic Potential of Medicinal Plants: A Review
Authors Info & Affiliations
Abstract
Introduction
Diabetes mellitus is a chronic metabolic condition characterized by hyperglycemia due to either insulin resistance, lack of insulin production, and/or secretion. The global rates of infection are increasing, especially in low- and middle-income countries; thus, exploring novel and safer therapeutic options has become essential. This review aims to compile and analyze existing data on the anti-diabetic properties of selected medicinal plants, focusing on their phytochemical composition, molecular mechanisms of action, and potential therapeutic roles in glucose homeostasis.
Methods
A comprehensive literature search was conducted using databases such as Scopus, ScienceDirect, and Google Scholar. Sixteen medicinal plants traditionally used for diabetes management were identified and examined based on multiple experimental studies.
Results
Phytochemicals such as flavonoids, polyphenols, saponins, and terpenoids from these plants exhibit anti-diabetic effects through multiple mechanisms: modulation of glucose transporters (e.g., GLUT4), activation of insulin signaling pathways (e.g., PI3K/Akt), inhibition of digestive enzymes (e.g., α-amylase, α-glucosidase), promotion of insulin secretion, and mitigation of oxidative stress.
Discussion
Notably, compounds like quercetin, kaempferol, berberine, and gymnemic acid have shown significant promise.
Conclusion
Medicinal plants present a rich source of bioactive compounds with multi-targeted mechanisms relevant to diabetes management. Further pharmacological investigations are necessary to establish their therapeutic efficacy and safety, enabling the development of phytochemical-based interventions for diabetes.
1. INTRODUCTION
Herbal therapy has been used for several thousand years and has gained increasing attention in recent years for its potential in managing many illnesses, and serves as a valuable basis for identifying new therapeutic agents [1, 2]. Several medicinal plants (various herbs, fruits, and grains) are reported for their anti-diabetic activity. This healing effect is due to the nature of phytochemicals present in them, a wide variety of secondary metabolites, including: alkaloids, glycosides, tannins, essential oils, and terpenoids; which in turn depend on the growing conditions, timing, and process of harvesting, and light and air exposure of the plant [3-5]. Diabetes is one of the most prevalent chronic Non-Communicable Diseases (NCDs) worldwide, the fourth leading cause of death after cardiovascular diseases, cancers, and chronic respiratory diseases [6]. In 2024, the IDF estimated that 11.1% (589 million) of adults worldwide were living with diabetes. Most of them (81%) are concentrated in developing countries, where limited access to diagnosis and proper treatment exacerbates the disease burden [7]. Diabetes requires early diagnosis, and its management includes lifestyle modifications, pharmacotherapy, and, in certain cases, insulin therapy [4]. Nonetheless, these conventional approaches are often linked to adverse effects, which has fueled growing interest in medicinal plants as alternative or complementary therapeutic options [8]. For more efficient and risk-free methods of reducing glucose, Numerous medicinal plants have shown promising glucose-lowering effects via several processes. They regulate blood glucose level through antioxidative effect, a crucial element in the onset and advancement of diabetes, through glucose and lipid metabolism regulation, and insulin resistance reduction [2, 9, 10]. This review provides an overview of medicinal plants with anti-diabetic properties. It focuses on phytochemicals identification, classification, and their biochemical and molecular mechanisms through which they exert the hypoglycemic effect.
2. METHODS
This review was conducted by performing a literature search of electronic databases: Scopus, Google Scholar, and ScienceDirect to collect relevant studies on the anti-diabetic potential of medicinal plants. Articles published between 2000 and 2025 were considered. The literature search used keywords such as “medicinal plants”, “anti-diabetic activity”, “phytochemicals”, “insulin signaling”, “α-amylase inhibition”, “α-glucosidase inhibition”, “in vitro”, “in vivo”, and “in silico”.
An initial search yielded 1,245 records. After removal of duplicates, 1,020 records were screened based on titles and abstracts. Selected papers were research articles or reviews that evaluated the anti-diabetic properties of medicinal plants or isolated phytochemicals. Only studies providing experimental evidence, such as in vitro, in vivo, or in silico approaches, were considered. These studies reported effects on glucose regulation, insulin signaling, enzyme inhibition (α-amylase, α-glucosidase), antioxidant activity, or related metabolic markers. Studies were excluded if they lacked experimental or mechanistic evidence, were not directly related to diabetes or metabolic dysfunction, or focused on synthetic drugs without plant-based components. Following full-text evaluation, 54 articles were included in the final review, covering 16 medicinal plants with reported molecular anti-diabetic mechanisms (Fig. 1).
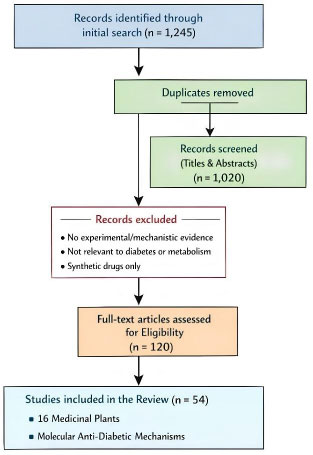
Literature search and selection process for the review.
2.1. Study Selection Process
Full texts of potentially eligible studies were assessed independently by the authors based on the predefined inclusion and exclusion criteria. From the included studies, a total of sixteen medicinal plants traditionally used for diabetes management were identified and analyzed.
3. RESULTS AND DISCUSSION
3.1. Medicinal Plants with Antidiabetic Potential
Based on the literature search carried out through the databases Scopus, ScienceDirect, and Google Scholar, 16 plant species used in traditional medicine with an antidiabetic effect were identified. The medicinal plants discussed below were selected based on the availability of experimental evidence supporting their molecular anti-diabetic effects. However, they differ in botanical origin and phytochemical composition. Many converge on common biological pathways relevant to glucose homeostasis, including inhibition of carbohydrate-digesting enzymes, modulation of insulin signaling, enhancement of glucose uptake, and attenuation of oxidative stress and inflammation. The medicinal plants reviewed in this section are summarized in Table 1, which compares their major bioactive compounds, principal antidiabetic mechanisms, experimental models, and strength of evidence. Selected plants are discussed below to highlight representative mechanisms and supporting studies.
| Medicinal Plant | Major Bioactive Compounds | Main Molecular Mechanisms Related to Diabetes | Experimental Model | Evidence Level | References |
|---|---|---|---|---|---|
| Arachis hypogaea (L.) (peanut shells) | Luteolin, pyrogallol, catechol, quercetin, polyphenols | Antioxidant activity; inhibition of α-amylase and carbohydrate digestion | In vitro | Limited | [11, 12] |
| Boswellia sacra (Flueck.) | Boswellic acid derivatives | Reduction of blood glucose; modulation of inflammatory and metabolic pathways | In vitro, in vivo & in silico | Moderate | [13, 14] |
| Berberis glaucocarpa (Stapf) | Berberine, palmatine, dihydroberberine derivatives | Hypoglycemic effects: improvement of glucose metabolism and insulin sensitivity | In vivo | Moderate | [15, 16] |
| Coccinia grandis (L.) Voigt | Cephalandrines, taraxerol, β-sitosterol, lupeol | Reduction of blood glucose; modulation of lipid and glucose metabolism | In vivo | Moderate | [17, 18] |
| Dittrichia viscosa (L.) | Polyphenols | Inhibition of α-glucosidase and α-amylase; increased GLP-1 levels; reduced glucose absorption | In vitro & in vivo | Moderate | [19, 20] |
| Gracilaria bursa-pastoris | Diterpenes, phenolic acids, bromophenols | Antioxidant and anti-inflammatory effects; indirect glucose-lowering activity | In vitro & in silico | Limited | [21, 22] |
| Gymnema sylvestre (Retz.) | Gymnemic acids, stigmasterol, quercitol | Stimulation of insulin secretion; β-cell protection; enhanced glucose uptake | In vitro & in silico | Strong | [23, 24] |
| Opilia acuminata Wall. ex Baill. | Kaempferol | Insulin secretion, increased cellular glucose uptake, and antioxidant effects | In vitro & in silico | Limited | [25, 26] |
| Xenostegia tridentata (L.) | Alkaloids, luteolin, and diosmetin derivatives | Hypoglycemic activity; modulation of carbohydrate metabolism | In vitro & in vivo | Moderate | [27, 28] |
| Morus alba (L.) | Rutin, quercetin-3-O-β-D-glucoside | Activation of AMPK; Akt-mediated insulin signaling; enhanced glucose uptake | In vitro, in vivo & in silico | Strong | [29, 30] |
| Phellinus igniarius (L.) | Styrylpyrones (phenolics) | Inhibition of α-glucosidase; suppression of protein glycation and oxidative stress | In vitro, in vivo & in silico | Moderate | [31–34] |
| Punica granatum (L.) | Phenols, tannins, triterpenoids | Reduction of blood glucose; antioxidant and anti-inflammatory mechanisms | In vitro & in vivo | Strong | [35, 36] |
| Red ginseng (Panax ginseng C.A. Meyer) | Ginsenosides (saponins), volatile oils | Improved insulin sensitivity; enhanced insulin signaling; increased fatty acid oxidation | In vitro, in vivo & in silico | Strong | [37, 38] |
| Saccharum officinarum (L.) | Tricin, apigenin, naringenin, and luteolin derivatives | Antioxidant and anti-inflammatory effects; indirect metabolic regulation | In vitro | Limited | [39, 40] |
| Vernonia amygdalina (Delile.) | Sesquiterpene lactones, flavonoids, saponins | Hypoglycemic and antioxidant effects. | In vivo | Moderate | [41, 42] |
| Wisteria sinensis (Sims) Sweet | Flavonoids (luteolin, apigenin derivatives), saponins | Activation of IRS-1/PI3K/Akt/GLUT4 pathway; improved glucose uptake | In vitro & in vivo | Moderate | [43-45] |
Arachis hypogea (L.) (Peanut shells) are abundant in polyphenols, luteolin, carotene, and iso-saponaretin [11]. Polyphenol-rich extracts from peanut shells exhibit antidiabetic activity primarily via antioxidant effects and inhibition of carbohydrate-hydrolyzing enzymes such as α-amylase; however, direct evidence linking these effects to insulin signaling or glucose uptake pathways remains limited. HPLC characterization revealed the presence of luteolin at the highest amount, followed by pyrogallol, catechol, phlorglucinol, and quercetin, which are responsible for the therapeutic effect [11, 12].
Boswellia sacra (Flueck.), which belongs to the Burseraceae family, generates aromatic oleo-gum resin, commonly referred to as frankincense, which traditionally stands out for medicinal purposes [13]. Studies reported active boswellic acid derivatives as the anti-diabetic agent of the plant, which lower blood glucose via several mechanisms of action [14].
Berberis glaucocarpa (Stapf), belonging to the Berberidaceae family, is widely distributed worldwide. Its root decoction is traditionally used for the treatment of diabetes [15]. Berberin, palmatine, and 8-trichloromethyl dihydroberberine are the plant's major compounds that confer its hypoglycemic activity [16].
Coccinia grandis (L.) Voigt belongs to the Cucurbitaceae family, native to the tropical areas of Asia, and plays a significant role in traditional medicine as a complementary therapy for diabetes mellitus management [17]. Phytochemical characterization revealed the presence of cephalandrine a, cephalandrine b, taraxerol, stigmast-7- en-3-one cephalandrol, β-sitosterol, lupeol, taraxerone, and tritriacontane, responsible for the hypoglycemic effect [18].
Dittrichia viscosa (L.) (Inula viscosa (L.) Aiton) is mainly distributed in Africa, Asia, and Europe. It is traditionally used for wound healing, rheumatic pain, and treatment of bronchitis and tuberculosis [19]. Its abundance in polyphenols contributes to anti-diabetic effects via: inhibition of glucose absorption in the intestine through inhibiting β-glucosidase and α-amylase activity, and increasing glucagon-like peptide 1 (GLP1) [20].
Gracilaria bursa-pastoris, a red macroalga primarily known for its culinary use rather than its medicinal properties, is distributed across temperate and tropical regions of the northeast Atlantic and the Mediterranean. [21]. It is composed of a variety of biologically active secondary metabolites: diterpenes, phenolic acids, sulfonic acids, bromophenols, heterosides, etc., which possess antidiabetic, antioxidant, and anti-inflammatory activities [22].
Gymnema sylvestre (Retz.), commonly known as “Gurmar,” belongs to the family Asclepiadaceae; it is one of the most popular anti-diabetic plants, distributed in Asia, tropical Africa, Malaysia, and Sri Lanka [23]. It exhibits glucose absorption modulation, stimulates insulin secretion, and promotes β-cell regeneration. Gymnema sylvestre represents one of the most extensively studied and mechanistically supported anti-diabetic medicinal plants [24].
Opilia acuminata Wall. ex Baill., belongs to the family Opiliaceae; it is cultivated and used to treat diabetes in Yunnan, Indonesia, Malaysia, Thailand, and Vietnam [25]. Numerous bioactive constituents were identified in it. Among them, kaempferol (its predominant compound) plays a key role by stimulating insulin release, promoting cellular glucose uptake, and mitigating oxidative stress [26].
Despite their diverse botanical origins, the reviewed species converge on a limited number of antidiabetic mechanisms, mainly insulin signaling modulation, inhibition of carbohydrate-digesting enzymes, and oxidative stress attenuation (Table 1). However, the strength of evidence differs substantially, with several plants supported mainly by in vitro or in silico data.
Xenostegia tridentata (L.) belongs to the family Convolvulaceae, is distributed in different parts of the world, and is used for the treatment of different disorders: swellings, rheumatism, urinary infections, and its root extracts are beneficial in the treatment of diabetes [27]. Several studies highlighted the abundance of bioactive components: alkaloids, luteolin-7-O-β-D-glucoside, diosmetin, diosmetin-7-O-β-D-glucoside, and luteolin, responsible for the anti-diabetic effect [28].
Morus alba (L.) (Moraceae), commonly known as the white mulberry tree, is native to Europe and Asia. Traditionally, it has been used to prevent various disorders and to help manage elevated blood glucose levels [29]. Phytochemical studies have identified two major compounds, rutin and quercetin-3-O-β-D-glucoside (Q3G), which enhance glucose uptake through the Akt-mediated insulin signaling pathway and activation of AMP-Activated Protein Kinase (AMPK) [30]. These findings place Morus alba among plants with robust molecular evidence targeting key regulators of glucose metabolism.
Phellinus igniarius (L.), commonly referred to as “Sanghuang”, has been used in oriental countries for the treatment of various diseases [31]. It is a fungus, well-known for its numerous therapeutic effects, including anti-diabetic and anti-inflammatory properties [33]. Characterization studies revealed that styrylpyrones (phenol components) from Phellinus species have multi-target anti-diabetic effects via: inhibition of α-glucosidase, protein glycation, and oxidative stress [32-34].
Punica granutam (L.) is a member of the Punicaceae family. It has been used in folk medicine for centuries in the Mediterranean region, the Middle East, India, and China [35]. Traditionally, it has been employed to alleviate conditions ranging from inflammation and rheumatism to mild throat pain. Its flowers have been used as an antidiabetic medicine as they can significantly lower the blood glucose level via different mechanisms due to their high content of phenols, tannins, and triterpenoids [36].
Red ginseng (steamed and dried form of Panax ginseng C.A. Meyer) is commonly used in Asian folk medicine for its various biological activities, especially antidiabetic properties [37]. It mainly contains saponins and volatile oils that improve insulin sensitivity, enhance insulin signal transduction, and promote fatty acid oxidation [38].
Saccharum officinarum (L.), known as sugarcane, native to south and southeastern Asia [39], is rich in phytochemicals such as flavonoids, stilbenes, phenolic acids, tannins, and lignans. Key phenolics, including tricin, apigenin, naringenin, and luteolin derivatives, exhibit antitumor, anti-inflammatory, antioxidant, and antihistamine activities [40].
Vernonia amygdalina (Delile.), belonging to the Asteraceae family, is one of the most popularly used in traditional medicine in Africa and Asia. Leaves and roots decoctions have been used to treat kidney problems, hiccups, fevers, stomach disorders, and diabetes [41]. Several studies reported active phytochemicals, including: terpenes, alkaloids, saponins, tannins, flavonoids, phenolics, steroidal glycosides, triterpenoids, and several types of sesquiterpene lactones that are responsible for the anti-diabetic activity [42].
Wisteria sinensis (Sims) Sweet (known as Chinese wisteria) belongs to the Leguminosae family and is widely abundant in Asian countries. It is commonly used in the treatment of stomach ulcers, cancers, and inflammatory rheumatism [43]. Its secondary metabolites, polyphenols, triterpenes, saponins, isoflavones, and lectins, give the plant strong antioxidant, antimicrobial, anti-inflammatory, and antitumoral properties [45]. The high content of flavonoids: luteolin-7-O-β-D-glucoside, apigenin 4′-O-β-D-glucopyranoside, apigenin-7-(6′′-acetyl-glucopyranoside), luteolin, and apigenin gives a beneficial hypoglycemic effect, alleviating type 2 diabetes via activation of IRS-1/PI3K/Akt/GLUT4 pathway [44].
Among the surveyed species, Gymnema sylvestre, Morus alba, Panax ginseng (red ginseng), and Punica granatum exhibit the strongest mechanistic support, supported by multiple in vitro and in vivo studies. Plants such as Dittrichia viscosa, Phellinus igniarius, and Wisteria sinensis display moderate mechanistic evidence. In contrast, species including Arachis hypogaea shells, Gracilaria bursa -pastoris, Opilia acuminata, and Saccharum officinarum are supported mainly by biochemical, antioxidant, or preliminary cellular assays, with limited direct validation of their molecular targets in diabetic models. However, contradictory findings regarding bioavailability, dose-dependence, and insulin secretagogue activity remain for several species, underscoring the need for standardized experimental designs and clinical validation. For most of the medicinal plants reviewed, clinical studies are currently lacking. Thus, the translational relevance of preclinical findings remains uncertain, and well-designed human trials are necessary.
Despite the apparent convergence toward common molecular targets, the strength of evidence varies considerably among the reviewed plants. Species classified as having strong evidence, such as Gymnema sylvestre, Morus alba, Panax ginseng, and Punica granatum, are supported by multiple in vivo studies and mechanistic investigations. In contrast, plants with limited evidence rely predominantly on in vitro enzymatic inhibition or antioxidant assays, which do not fully reflect the complexity of glucose homeostasis in vivo. Moreover, differences in extraction procedures, phytochemical standardization, and experimental models limit direct comparison between studies and may partly explain discrepancies in reported efficacy. The convergence of anti-diabetic mechanisms observed among the reviewed medicinal plants can be largely attributed to a set of recurrent bioactive phytochemicals. Identifying these compounds and elucidating their molecular actions is essential. Thus, attention is next directed toward the phytochemicals responsible for anti-diabetic effects. The subsequent subsection discusses their molecular mechanisms of action in diabetes mellitus.
3.2. Bioactive Phytochemicals' Effects on Diabetes Mellitus
Diabetes Mellitus (DM) is characterized by increased levels of blood sugar (glucose) above the physiological range, due to insufficient insulin activity and/ or secretion [28]. Its etiology varies, leading to different types: type 1 diabetes (an autoimmune condition causing beta-cell death), type 2 diabetes (the most prevalent type characterized by insulin resistance), gestational diabetes (may be developed during pregnancy), and other types of diabetes (caused by genetic defects in beta-cell function, pancreatic disorders, or associated with endocrinopathies) [3, 6]. Many plants have been used as folk medicine to ameliorate diabetes patients’ conditions, without side effects and via several mechanisms of action [46]. Various plant compounds and their molecular antidiabetic effects are reported in this part.
3.2.1. GLUT Transport and Insulin Pathway Modulation
Numerous studies have been conducted on the modulation of glucose transporters (GLUT) activities, transmembrane proteins responsible for glucose uptake via insulin-stimulated transport into skeletal muscle and adipose tissue [47, 48]. An in vivo results study reported a great hypoglycemic activity of Wisteria sinensis flowers’ extract, lowering blood glucose [44]. The mechanism of action was explored by an in vitro investigation. Western blot results showed that this extract significantly decreased IRS-1 phosphorylation, enhancing PI3K expression, elevating Akt phosphorylation and GLUT4 expression and its translocation into the plasma membrane of L6 cells, inducing glucose uptake in liver and skeletal muscle, thus contributing to hyperglycemia reduction [44]. GLUT1 expression was also enhanced in both the transcription and translation levels in the presence of Chimonanthus nitens leaf extract [49]. A relevant study showed the hypoglycemic effect of Peanut Shell Polyphenol Extract (PSPE). The treatment reversed the defective glycogen storage in the diabetic state, probably via the promotion of GLUT1 for glucose absorption into the hepatic tissues and its conversion into glycogen [12]. Another in vivo study on a type 2 diabetic mouse model demonstrated the anti-diabetic activity of Phellinus igniarius (L.) polyphenol-rich extract (PI-PRE), which displayed, after a 4-week treatment, a stimulatory effect on GLUT4 expression and translocation, alongside this, p-AMPKα/AMPK ratio up-regulation, analyzed by western blotting, leading to glucose uptake [50]. A different investigation also revealed the modulation effect of Punica granatum flower polyphenols extract (PGF) on the insulin pathway. Western blotting of markers from the liver tissue obtained after the in vivo study showed enhanced phosphorylation of IR, Akt, and GSK-3b, leading to improved insulin signaling activity, thus, to glucose uptake and an increase of glycogen content [35]. A further study provided evidence on the anti-diabetic effect of L. sylvestris extract evaluated in vitro, where isolated compounds: Interruptin A, interruptin C, ergosterol, and diglycerol promoted insulin secretion from beta cells of the INS-1E cell line [26]. A similar investigation on the effect of Polyphenol-Rich Sugarcane Extract (PRSE) on glucose uptake in the Caco-2 cells line (which mimics the human intestinal cells in sugar absorption capacity) demonstrated an inhibition of intestinal glucose absorption by affecting both facilitated diffusion and active transport mechanisms, also inhibiting cellular production of GLUT2, thus, intestinal sugar uptake [39]. Similar findings were reported by another study [18]. New evidence from another investigation reported the antidiabetic effect of rutin and Q3G isolated from the fruits of Morus alba, on 3T3-L1 adipocytes cell line, results revealed that these two compounds improved glucose uptake by a simultaneous activation of AMPK (Akt phosphorylation) and the insulin signaling pathway [30]. Fig 2 below summarizes these activities. While activation of the PI3K/Akt/GLUT axis appears as a recurrent mechanism across multiple plant extracts, most evidence is derived from cell-based models such as L6, 3T3-L1, and HepG2 cells. These systems do not fully reproduce insulin resistance or metabolic regulation observed in human type 2 diabetes. Furthermore, differences in extract composition and concentrations used across studies make it difficult to determine whether the observed effects result from direct insulin mimetic activity or secondary metabolic adaptations.
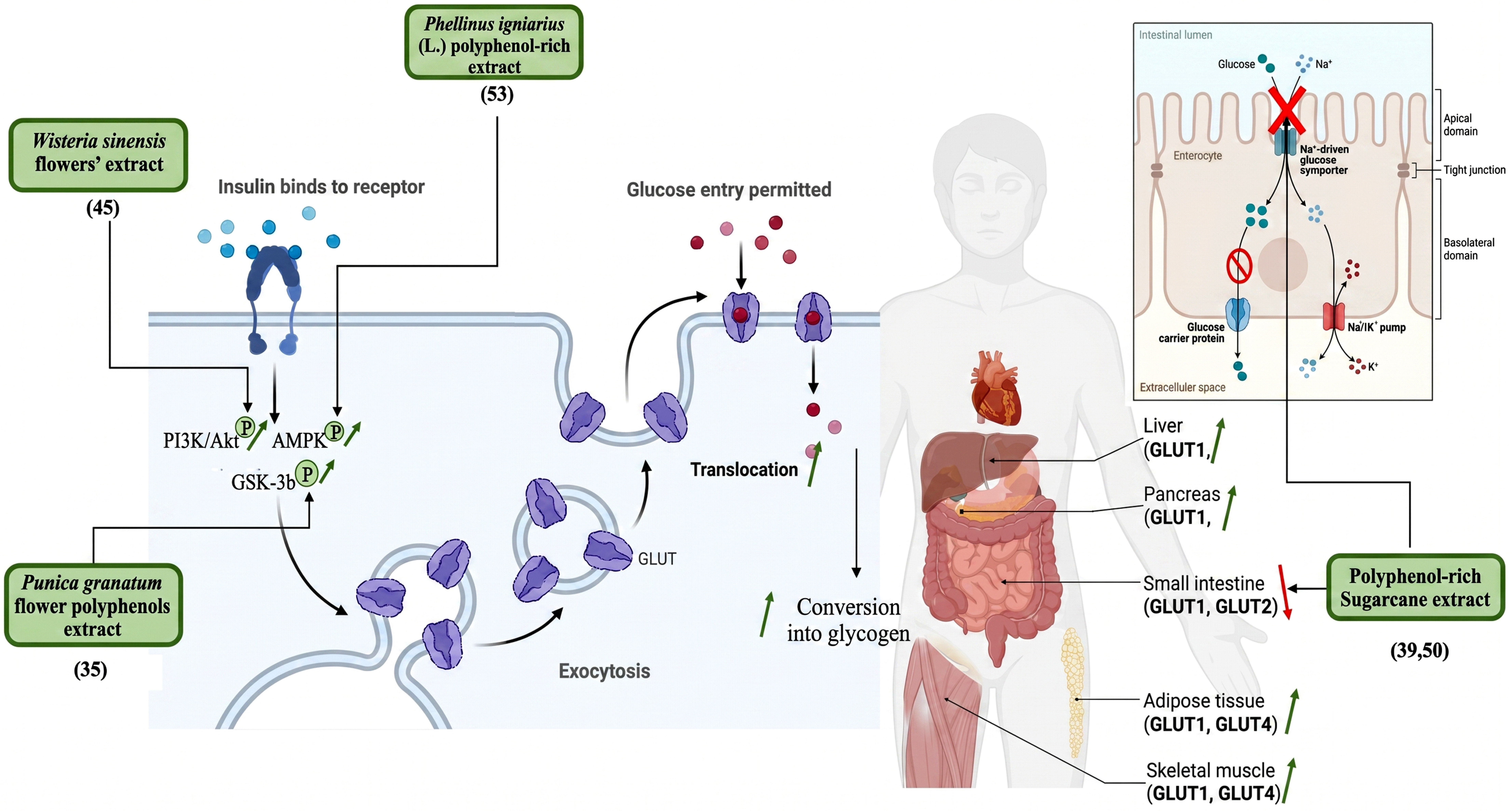
Insulin and GLUT pathways modulation of different medicinal plant extracts. It illustrates the modulation of insulin signaling and glucose transport by polyphenol-rich extracts from Wisteria sinensis, Punica granatum, Phellinus igniarius, and Saccharum officinarum. AMPK and PI3K/Akt pathways are consistently activated across multiple plants, leading to GLUT translocation and enhanced glucose uptake in liver, muscle, and adipose tissue.
3.2.2. Blood Glucose Level Modulation
Several studies explored the modulating effect of plant extracts on blood glucose release and uptake. A relevant study demonstrated in vitro and in silico potent inhibitory activity of quercetin and kaempferol, identified from Gymnema sylvestre leaves extract, against α-amylase enzyme. Additionally, a stable and precise interaction of rutin within the active site of peroxisome proliferator-activated receptor-gamma (PPARγ) protein, exhibiting an agonist effect, which improves the glucose-sensing abilities of pancreatic β-cells and insulin sensitivity, thus enhancing glucose uptake [24]. The hypoglycemic effect of different Gracilaria bursa-pastoris extracts (hexane, ethyl acetate, methanol, and water extracts) was revealed via in vitro inhibitory activities on pancreatic α-amylase and α-glucosidase enzymes. Molecular docking showed that rutin formed a stable and favorable interaction with the α-amylase active site [21]. Another in vivo study conducted on alloxan-induced diabetic mice showed that the flavonoids: cymaroside and cosmosiin present in Xenostegia tridentata (L.) stem and roots’ water and ethanol extracts have strong hypoglycemic activities. In vitro assay proved that the extracts exhibited inhibitory effects on key enzymes (α-amylase and α-glucosidase) that play an important role in hydrolyzing glycosidic bonds [28]. The in-silico part revealed that cymaroside and cosmosiin had the greatest binding affinity with α-amylase and α-glucosidase, thus, helping in the decrease of glucose absorption in the intestines, delaying its liberation into the bloodstream. They also had the best dock conformation with glycogen phosphorylase (GP) receptor, inducing glycogenolysis inhibition, thus, reducing hepatic glucose production and lowering blood glucose; moreover, quercetin had the best binding affinity with PPAR-γ, it established four bonds with ASP260, and SER342 amino acids. Thus, could facilitate fatty acid uptake into adipocytes and the transfer of lipids from non-adipose to adipose tissues [28]. The anti-α-glucosidase activity of Opilia acuminata leaf extract was confirmed through in vitro and in silico studies. Interruptin C and the novel diterpene compounds uncompetitively bind to HIS 280 in the enzyme’s catalytic pocket, inhibiting its activity [26]. However, inhibition of α-amylase and α-glucosidase, while effective in reducing postprandial hyperglycemia, is associated with gastrointestinal side effects and may offer limited benefit as a standalone strategy. Moreover, many conclusions in this category are based on molecular docking and in vitro enzyme inhibition, which do not account for bioavailability, intestinal metabolism, or compound stability under physiological conditions [24]. Other studies also showed the in vivo anti-diabetic effect of Peanut shell polyphenol extract (PSPE) and Punica granatum flower polyphenols extract (PGF). The oral glucose tolerance test results indicated that after PSPE and PGF treatments, an improvement in glucose intolerance in diabetic rats was observed by decreasing blood glucose compared to the diabetic control group [12, 35]. A different study reported the effect of Polyphenol-Rich Sugarcane Extract (PRSE) on the restoration of insulin production in dysfunctional β-cells. The in vitro treatment of glucotoxic-induced insulin-dysfunctional rat pancreatic β-cell model was able to restore insulin production by 39.48%, which is a great approach for type 2 diabetic treatment [39]. Another in vivo study demonstrated the anti-diabetic effect of Dittrichia viscosa on STZ-induced diabetes in rats. Besides lowering blood sugar measured by a colorimetric method, phosphoenolpyruvate carboxykinase (PEPCK), a crucial enzyme at the early step of the gluconeogenic pathway, was also reduced when treated with gold nanoparticles of D. viscosa (AuNPs). Even its mRNA expression levels were decreased, leading to insulin secretion enhancement, thus to glucose uptake and hypoglycemic effect [20]. All these therapeutic activities are regrouped in Fig. (3).
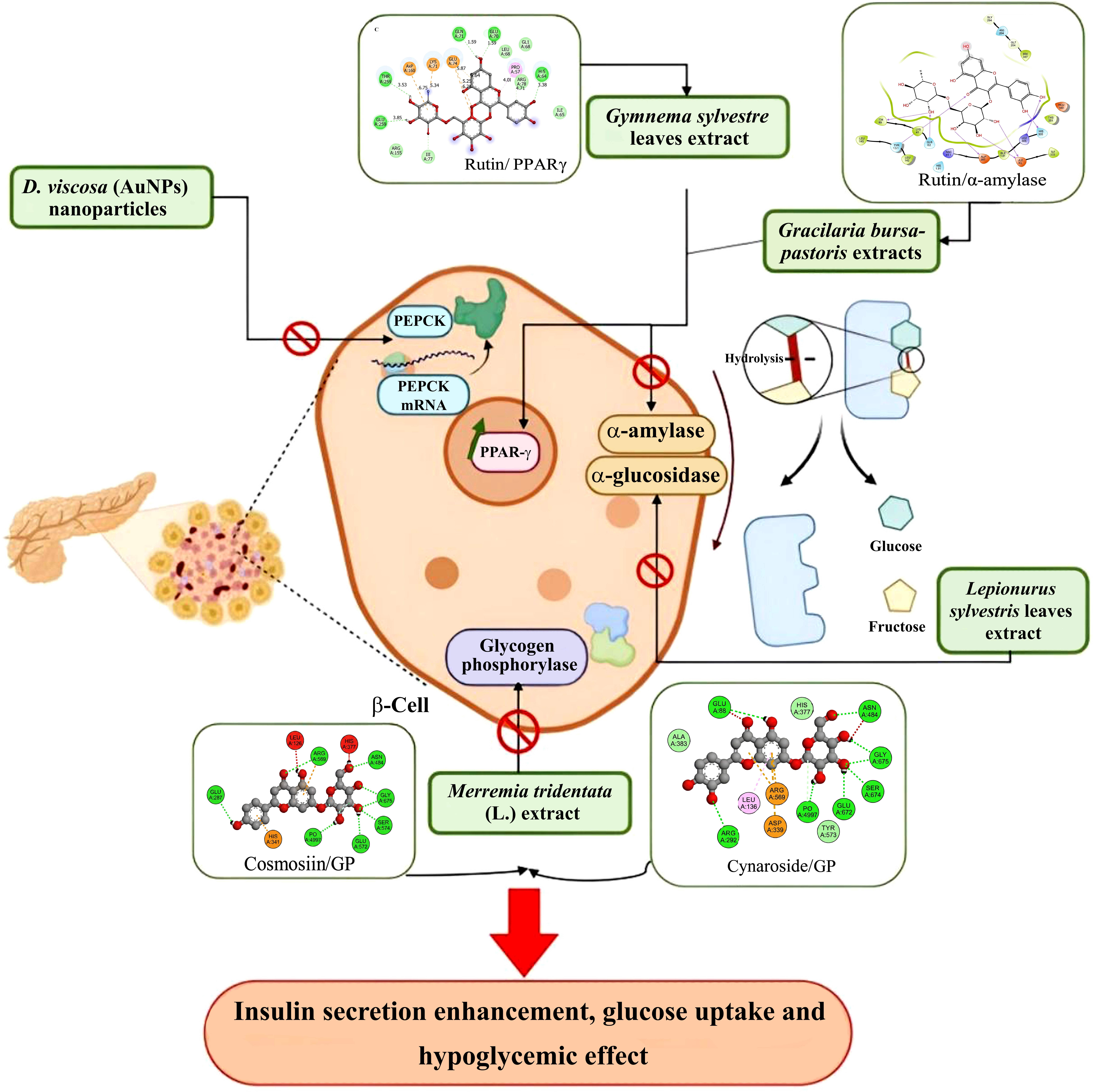
Phytochemicals' potential on carbohydrate digestion enzymes and gluconeogenesis inhibition. This figure summarizes the effect of Gymnema sylvestre, Dittrichia viscosa, Xenostegia tridentata, Opilia acuminata, and Gracilaria bursa-pastoris extracts on insulin secretion, α-amylase/α-glucosidase inhibition, and glycogen metabolism. Activation of PPAR-γ, inhibition of α-amylase/α-glucosidase, and glycogen phosphorylase suppression are observed across several species, promoting insulin secretion and lowering blood glucose.
3.2.3. Reactive Oxygen Species (ROS) Production Modulation
The pathophysiology of diabetes has been linked to oxidative stress either as a causative factor or because of its consequences; it contributes to cellular structural abnormality of the liver, kidney, and pancreas [51]. An in vivo investigation showed that pomegranate (Punica granatum L.) peel extract (PPE), combined with L-carnitine, treated mice for 12 weeks, restored the antioxidant enzyme “nuclear factor erythroid 2-related factor 2” (Nrf2) activity after induced diabetes. It also enhanced the activity of antioxidant enzymes (catalase (CAT) and superoxide dismutase (SOD)), thus downregulated the hyperglycemia through the antioxidant system [52]. An additional in vivo study supported the anti-hyperglycemic effects of PSPE via antioxidant activity, restoring SOD, Glutathione (GSH), and Malondialdehyde (MDA) hepatic levels, thus reversing pathological changes in the pancreatic islets, improving cellular structures [12]. Similar in vivo results of CAT, SOD, and glutathione peroxidase (GPx) levels restoration were revealed, supporting the anti-diabetic effect of PGF via oxidative stress regulation [35]. Consistent with the mechanisms described above, the polyphenol-rich Vernonia amygdalina leaves extract expressed similar results, where pancreatic SOD, CAT, and GPx activities were enhanced. In addition, GSH level was restored [42]. This was also seen in another investigation, which examined the in vivo antidiabetic effects of palmatine, berberine, and 8-trichloromethyl dihydroberberine derived from Berberis glaucocarpa. It enhanced SOD and CAT antioxidant enzyme levels [16]. Another subsequent research result revealed that red ginseng extract regulates oxidative stress, via an upregulation of LysoPCs and LysoPEs phospholipids (metabolites of glycerol-phospholipids), regulating the metabolism disorder, the oxidative state, and improving islet tissue damage [38]. An additional report on the anti-diabetic effect of β-Boswellic acid (β-BA) and 11-keto-β-boswellic acid (β-KBA) isolated from B. sacra, in vivo treatment on streptozotocin (STZ) -induced diabetic rats showed a restoration of oxidative state balance. It reduced plasma MDA (inhibiting lipid peroxidation) and increased SOD antioxidant enzyme activity. Alongside this, it demonstrated a reversal of the pathological alterations in pancreas and kidney histological architectures [14]. These effects are summarized in Fig. (4). Although antioxidant activity is consistently reported across many plants, oxidative stress modulation alone may not be sufficient to achieve sustained glycemic control. In several studies, improvements in antioxidant markers were not systematically correlated with insulin sensitivity or glucose uptake, highlighting the need to distinguish between supportive cytoprotective effects and direct antidiabetic mechanisms.
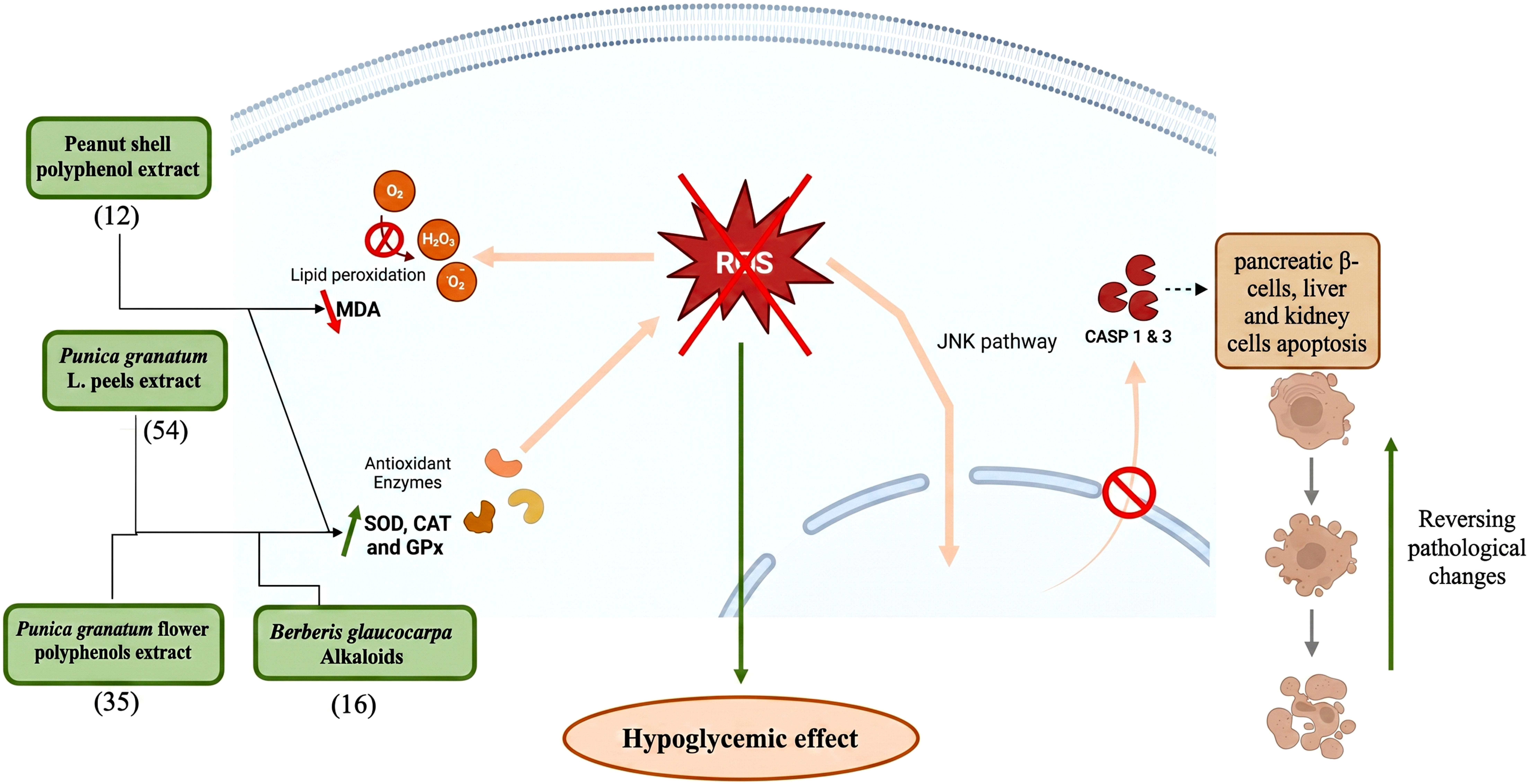
Key roles of phytochemicals from different medicinal plants in ROS scavenging activities for anti-diabetic effect. It illustrates the antioxidant and cytoprotective mechanisms of Arachis hypogaea, Punica granatum, and Berberis glaucocarpa extracts. These plants reduce ROS and lipid peroxidation, enhance SOD, CAT, and GPx activity, and protect pancreatic β-cells, liver, and kidney from apoptosis. ROS scavenging and antioxidant enzyme activation appear as consistent protective mechanisms across these species.
It is important to note the limitations in translational relevance. The majority of evidence is derived from in vitro, in vivo, or in silico studies, and no clinical trials are available to confirm efficacy in humans. Furthermore, factors such as bioavailability, pharmacokinetics, and metabolic stability of phytochemicals can significantly influence therapeutic outcomes. Effective concentrations observed in experimental studies may not be achievable through conventional dosing of plant extracts. In addition, variability in plant composition due to species differences, extraction methods, and environmental conditions may affect the reproducibility and reliability of results. These limitations underscore the need for carefully designed clinical studies and standardization of extracts before medicinal plants or isolated phytochemicals can be reliably translated into safe and effective anti-diabetic therapies. Additionally, future studies should integrate standardized extract characterization, dose–response analyses, and combined pharmacokinetic–pharmacodynamic evaluations to better correlate molecular mechanisms with therapeutic efficacy.
CONCLUSION
This review highlights the molecular anti-diabetic potential of sixteen medicinal plants traditionally used for diabetes management, demonstrating that their efficacy largely converges on a limited number of core biological pathways. Across the surveyed literature, plant-derived phytochemicals, particularly flavonoids, alkaloids, and polyphenols, consistently modulate insulin signaling cascades, including IRS-1/PI3K/Akt and AMPK pathways, promote GLUT4-mediated glucose uptake, inhibit carbohydrate-digesting enzymes (α-amylase and α-glucosidase), and attenuate oxidative stress. Collectively, these mechanisms support their hypoglycemic effects and their relevance to type 2 diabetes pathophysiology.
Despite this promising mechanistic evidence, most data remain derived from in vitro, in vivo, or in silico studies. Important translational challenges persist, including limited information on bioavailability, dose-response relationships, long-term safety, and variability in phytochemical composition among crude extracts. In addition, discrepancies in experimental models and methodologies complicate direct comparison across studies and hinder clinical extrapolation.
Future research should prioritize the standardization of plant extracts, detailed pharmacokinetic and toxicological evaluation of key bioactive compounds, and well-designed clinical trials to validate therapeutic efficacy. Addressing these gaps will be essential for translating molecular insights into evidence-based, plant-derived interventions that may serve as effective adjuncts or alternatives in diabetes management.
AUTHORS’ CONTRIBUTIONS
The authors confirm contribution to the paper as follows: S.O.: Article analysis and drafting of the original manuscript; M.T.: Conceptualization and drafting of the original manuscript; N.Be., L.A. and N.Br.: Supervised the writing and editing process. All authors read and approved the final manuscript.
LIST OF ABBREVIATIONS
| AMPK | = AMP-activated protein kinase |
| Akt | = protein kinase B |
| AuNPs | = gold nanoparticles |
| CAT | = catalase |
| GLUT | = glucose transporter |
| GLUT2 | = glucose transporter type 2 |
| GLUT4 | = glucose transporter type 4 |
| GP | = glycogen phosphorylase |
| GPx | = glutathione peroxidase |
| GLP-1 | = glucagon-like peptide-1 |
| GSH | = glutathione |
| GSK-3β | = glycogen synthase kinase-3 beta |
| INS-1E | = rat pancreatic beta-cell line |
| IR | = insulin receptor |
| IRS-1 | = insulin receptor substrate-1 |
| L6 | = rat skeletal muscle cell line |
| LysoPCs | = lysophosphatidyl cholines |
| LysoPEs | = lysophosphatidyl ethanol amines |
| MDA | = malondialdehyde |
| Nrf2 | = nuclear factor erythroid 2-related factor 2 |
| PEPCK | = phosphoenolpyruvate carboxykinase |
| PGF | = Punica granatum flower polyphenols extract |
| PI3K | = phosphoinositide 3-kinase |
| PPAR-γ | = peroxisome proliferator-activated receptor gamma |
| PRSE | = polyphenol-rich sugarcane extract |
| PSPE | = peanut shell polyphenol extract |
| Q3G | = quercetin-3-O-β-D-glucoside |
| ROS | = reactive oxygen species |
| SOD | = superoxide dismutase |
| STZ | = streptozotocin |
| β-BA | = β-boswellic acid |
| β-KBA | = 11-keto-β-boswellic acid |
CONFLICT OF INTEREST
Maria Trapali is an Associate Editorial Board Member of The Open Biochemistry Journal.
ACKNOWLEDGEMENTS
Declared none.

